Схема расключения или соединения электрических кабелей в распределительной коробке
Кабели и провода в жилых и нежилых строениях проложены от щита со счетчиком расхода электроэнергии, предохранительными пробками или автоматическими выключателями под полом, по стенам до распределительных коробок, где они соединены скручиванием или с помощью клеммников (электрических зажимов).
От раcпредкоробок электрическую проводку прокладывают к другим распаячным или расключительным коробкам, которые подключают на один автомат или одну и ту же группу и к выключателям, розеткам, светильникам. Выполнить расключение возможно и без специального образования и опыта работы электриком, достаточно следовать данной инструкции.
Обычно в квартиру приходят 3 – 4 группы (включая отдельную группу, приходящую на электроплиту, которую прокладывают независимыми кабелями или проводами). Таким образом, при повреждении на линии от электроснабжения отключается только часть строения или квартиры. Обратить особое внимание на пропорциональное и равномерное распределение нагрузки между автоматами или предохранительными пробками в электрическом щите.
До начала работ необходимо обесточить линию, на которой будут производиться работы, вывинтив пробки или отключив автомат.
Как правило, устанавливают одну распределительную коробку на каждое помещение, в случае значительного отдаления розеток может быть установлена дополнительная, специально на розетки.
Пошаговое руководство по монтажу распредкоробки
Подробнее о выборе способа соединения и особенностях монтажа на видео
Схема расключения распределительной коробки
Все защитные или заземляющие (на схеме маркированы как PE, выделены желтым цветом) проводники и нулевые (на рисунке маркированы N, обозначены синим) соединяют между собой по цветам, как показано на схеме. Проводника“земля” не будет, если электропроводка двухпроводная.
Проводника“земля” не будет, если электропроводка двухпроводная.
Фазные проводники (выделены черным и красным цветом) расключаются немного сложнее, если от распредкоробки проводка идет только к розеткам, то фазы также соединяют между собой.
В случае когда проводка идет на светильник с одноклавишным выключателем (как изображено на рисунке), провод, который отходит от выключателя, соединить с фазой, идущей на светильник (на рисунке L на освещение), а фазу, приходящую на выключатель, соединитьсо всеми фазными проводами (на схеме L на выключатель). Должно быть 4 соединения.
При распределении проводников на светильник с двухклавишным выключателем до него прокладывают четырехжильный кабель (2 фазных жилы включения, 1 фазная выключения,1 нулевая, 1 “ земля”). В случае двухпроводной электропроводки–трехжильный, так как проводник “земля” отсутствует. Соединяют между собой фазные проводники, идущие на розетки. Также соединяют фазы питающего кабеля, приходящие на общий зажим двухклавишного выключателя, 2 жилы с которого идут на лампы светильника отдельно. Ниже представленыс хемы распайки двухклавишного переключателя без “земли”.
Ниже представленыс хемы распайки двухклавишного переключателя без “земли”.
Все нулевые и заземляющие проводники (при наличии последних) соединяют вместе. Внутри коробки должно получиться 5 соединений (включая соединение заземляющих проводников). Ниже представлена схема расключения двухклавишного переключателя с“землей”.
Внимание! Перед подачей напряжения ещё раз внимательно проверить подключения. Во избежание короткого замыкания все соединения должны быть выполнены строго по схеме, изоляция должна быть неповрежденной, места скруток надежно изолированы или защищены изолирующими колпачками.
Схема соединения проводов в распределительной коробке
При самостоятельном ремонте электропроводки следует уделить особое внимание соединению проводов в распределительных коробках, ведь электротехника, как говорят электрики, это наука о контактах, как желательных, обеспечивающих бесперебойное снабжение электроэнергией, так и неприемлемых, из-за которых случается множество аварий и несчастных случаев.
В данной статье при помощи ссылок на другие материалы данного ресурса будет представлен весь багаж минимальных знаний и навыков, необходимый домашнему мастеру для успешного электромонтажа, а также будет поэтапно описан весь процесс соединений проводов в распределительной коробке, начиная от подготовительных этапов, заканчивая введением смонтированной домашней сети в эксплуатацию.
Схема – самый главный этап
Для бывалого электрика не составит труда подключить выключатель и лампочку без чертежа, сделав соединение проводов в распределительной коробке, ориентируясь по цветовой маркировке жил кабелей или по результатам прозвонки уже проложенной проводки.
Но у опытных мастеров такая работа получается лишь потому, что схема подключения отчетливо отпечаталась в уме благодаря многолетней практике.
Простая схема соединений проводов в распределительной коробке при подключении выключателя и лампочкиЕсли же мастеру требуется провести электропроводку на любом объекте, то он начнет первым делом с проектировки схемы, если до него это не сделали другие специалисты. Разумеется, существуют нормативы, ГОСТ и СНиП, по которым делаются все чертежи электропроводки, но для квартиры или частного дома подойдет схема, начертанная от руки, главное – чтобы она впоследствии исправно и безопасно работала.
Разумеется, существуют нормативы, ГОСТ и СНиП, по которым делаются все чертежи электропроводки, но для квартиры или частного дома подойдет схема, начертанная от руки, главное – чтобы она впоследствии исправно и безопасно работала.
Такой подход не только страхует от ошибок, но и позволяет экономить время, разделяя труд – осуществляя электромонтаж в распределительной коробке, электрик не будет отвлекаться на составления плана электропроводки, одновременно пытаясь удержать в голове планируемые линии и подключения.
Более сложная схема подключения двухклавишного выключателяПримеры схем для однокомнатной и двухкомнатной квартиры можно посмотреть, перейдя по ссылкам. Также нужно будет ознакомиться с условными графическими обозначениями элементов сети, чтобы можно было проконсультироваться у знакомого мастера, показав ему схему, или в будущем осуществить ремонт электропроводки, сверившись с сохраненным чертежом.
Важность планирования электропроводки обуславливается также и рациональностью и экономичностью. При составлении схемы проводки нужно составить план таким образом, чтобы самих распределительных коробок в квартире было как можно меньше, ведь каждое соединение проводов увеличивает риск потери контакта.
При составлении схемы проводки нужно составить план таким образом, чтобы самих распределительных коробок в квартире было как можно меньше, ведь каждое соединение проводов увеличивает риск потери контакта.
При подключении одной розетки или выключателя многие электрики советуют вообще обойтись без распределительной коробки и осуществить подключение проводов в подрозетнике. Такое решение обуславливается нежеланием владельцев квартир обеспечивать доступ к распределительной коробке, как того требует ПУЭ (по их мнению портится интерьер жилища). Розетка или выключатель легко разбирается, что обеспечивает доступ к соединению проводов в подрозетнике.
Подробный рассказ о различных схемах подключения проводов в распределительных коробках показан на видео:
Правильная маркировка выводов
Для того чтобы осуществить соединение проводов в распределительной коробке согласно схемы без излишних сомнений, нужно, чтобы выводы проложенной электропроводки были промаркированы.
Чтобы не тратить время на прозвонку кабелей, которая также описана на данном сайте, нужно еще на этапе укладки кабелей маркировать их окончания. В некоторых случаях, если проводов в распределительной коробке мало, можно ориентироваться по цвету изоляции токонесущих жил. Но при большом скоплении кабелей цвета изоляции проводов будут повторяться, поэтому возрастает вероятность ошибки.
Для исключения возможности неправильного подключения необходимо маркировать окончания уложенных кабелей любым доступным способом. Промышленность выпускает множество различных бирок, предназначенных для маркировки выводов. Очень часто электрики используют прозрачные термоусадочные трубки, вставляя полоску бумаги с условным обозначением вывода вовнутрь.
Также можно воспользоваться прозрачным скотчем, обматывая им бирки с надписями. В крайнем случае, маркером или фломастером написать обозначения прямо на изоляции проводов или на стене, — но при этом есть риск, что надписи сотрутся в процессе монтажа.
Маркировка выводов электропроводки произведена на стенеСледует также неуклонно придерживаться цветовой маркировки кабелей (коричневый, красный, черный – фаза, синий – рабочий ноль, зеленый с желтыми полосками – защитный проводник PE).
Общепринятая цветовая маркировка изоляции кабельных жилМонтаж распределительной коробки
Перед началом электромонтажа нужно убедиться, что распределительная коробка надежно зафиксирована. Виды распределительных коробок, которые зависят от типа электропроводки, также описаны на данном сайте. Важность фиксации обуславливается возможностью повреждения уже соединенных проводов при манипуляциях с распределительной коробкой и кабелями.
При работе с одножильными проводами следует избегать частых изгибаний токонесущих жил. Многожильные провода хоть и допускают частые изгибы, но механические нагрузки и вес незакрепленного кабеля висящей электропроводки может повредить уже готовое соединение, поэтому важно не нарушать технологию электромонтажа.
Если устанавливается скрытая проводка в кирпичной или бетонной стене, то провода укладываются в штрабу, проделываемую вручную или при помощи инструментов (штробление), а подрозетники и распределительные коробки монтируются в отверстиях, которые сверлятся в стене при помощи специальной коронки. Фиксация осуществляется при помощи гипса или алебастра.
Коронка для сверления отверстий для распределительных коробокКаждый тип соединения проводов требует определенной длины выводов. Например, при использовании весьма громоздких соединительных колодок, на которых имеются болтовые клеммы подключения, мастера пытаются оставлять свободной длины проводов как можно меньше, чтобы все соединения поместились в коробку.
Но, если используется сварка проводов, то длина оголенных проводников должна быть около семи сантиметров, чтобы не допустить оплавления изоляции при сильном нагреве, плюс длина изолированных жил для удобного и безопасного монтажа. Длинные оголенные соединенные проводки изолируются и укладываются в распределительной коробке.
Соединения проводов выполнены с помощью сваркиПриведенные выше примеры означают, что независимо от выбранного способа соединения проводов, длина выводов, выступающих из распределительной коробки, должна быть не менее десяти сантиметров, ведь излишки проводников всегда можно обрезать, но нарастить их будет чрезвычайно трудно, и при этом ухудшится надежность всей линии электропроводки.
Подготовка проводников
На данном этапе мастер должен окончательно решить, как соединить провода в распределительной коробке, и соответственно подготовить проводники – снять изоляцию и подогнуть жилы кабелей. Перечень существующих типов соединений проводов приводится ниже, а сейчас, независимо от выбранного способа, следует уделить особое внимание качеству зачистки и подготовки проводников к электромонтажу.
Для проведения любого электромонтажа мастер должен иметь набор инструментов электрика, ознакомиться с которым, можно, перейдя по ссылке. На данном этапе производится подготовка проводов к подключению – зачистка от изоляции. Следует с особой тщательностью отнестись к данному процессу, так как при использовании подручных средств (ножа, плоскогубцев, кусачек) возможны незаметные повреждения токонесущих жил.
Профессиональный набор инструментов электромонтажникаПри изгибании проводов и последующем монтаже едва заметные трещинки в металле могут увеличиться, что ухудшит электрическую проводимость и может привести к разлому и потере контакта. При больших токах в самом тонком участке цепи (в месте трещины) будет выделяться большое количество тепла, которое может разогреть изоляцию и даже сам металл проводника до температуры плавления.
При большом токе в надломанном проводнике происходит лавинообразный процесс – при нагревании сопротивление материала возрастает, что еще больше увеличивает тепловыделение.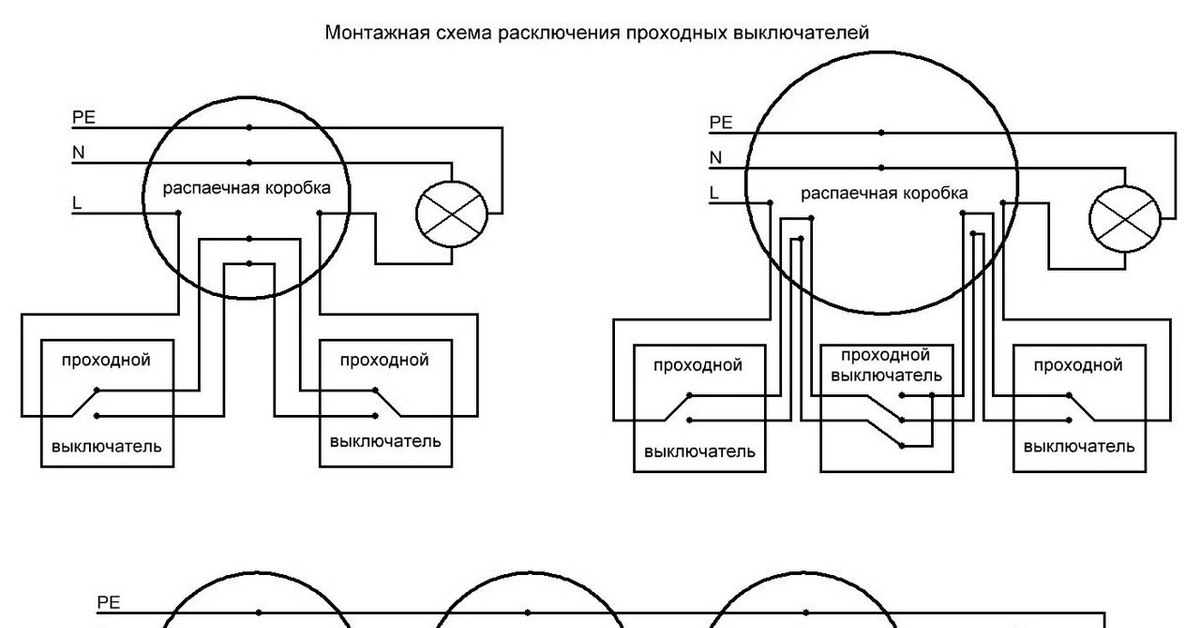 Заканчивается данный процесс расплавлением металла проводника и возникновением электрической дуги, которая обжигая другие соединения проводов в распределительной коробке, приводит их в полную негодность.
Заканчивается данный процесс расплавлением металла проводника и возникновением электрической дуги, которая обжигая другие соединения проводов в распределительной коробке, приводит их в полную негодность.
Без сомнения, настоящий мастер даже с помощью кухонного ножа сможет аккуратно и быстро снять изоляцию без повреждения токонесущей жилы. Но людям без опыта будет очень трудно избежать малозаметных повреждений материала проводников, поэтому, чтобы соединение проводов в распределительной коробке было качественным, необходимо использовать специальные инструменты для снятия изоляции (стрипперы).
Разнообразие инструментов для снятия изоляцииМногим доводилось наблюдать картину, что некоторые мастера снимают изоляцию при помощи зубов. Абсолютно не стоит следовать такому примеру, и в этом случае покупка профессионального инструмента для снятия изоляции обойдется дешевле, чем услуги стоматолога.
Обзор различных инструментов для снятия изоляции показан на видео:
 youtube.com/embed/HWsIEUPCNc8″ frameborder=»0″ allowfullscreen=»allowfullscreen»/>
youtube.com/embed/HWsIEUPCNc8″ frameborder=»0″ allowfullscreen=»allowfullscreen»/>Способы соединения проводов
Как уже говорилось, для различных способов соединений проводов распределительной коробке существует своя технология электромонтажа, которая заключается в длине зачищенных проводников, их изгибании и использовании соответствующих инструментов.
Поскольку каждый из возможных способов имеет множество специфических нюансов и требует применения специальных инструментов и навыков, ниже, в виде списка со ссылками представлены типы соединений проводов:
Электромонтаж проводов в распределительной коробке при помощи клеммных колодокКаждый из приведенных выше способов имеет свои достоинства и недостатки, ознакомиться с которыми нужно обязательно, изучив материалы, перейдя по предложенным ссылкам. Выбирать тип соединений для проведения электромонтажа своими руками следует исходя из наличия инструментов, навыков, возможности приобретения материалов и ожидаемого качества контактов.
Наиболее надежной считается сварка проводов, но она требует специального сварочного аппарата и специфических навыков. Пайка проводников, которая также имеет хорошие характеристики, требует навыков владения паяльником. Клеммники Wago достаточно просты в монтаже, надежны при правильно подобранных изделиях согласно нагрузке, допускают соединение многожильных проводов без использования специальных наконечников, но следует опасаться подделок.
Соединения в распределительной коробке выполнены при помощи клеммников WagoИспользование гильз надежное, при применении специальных изделий можно соединять медь и алюминий, но само соединение требует специальных клещей и является неразборным, что не дает шанса на легкое исправление ошибки электромонтажа. Колпачки СИЗ надежны при соблюдении технологии и правильном подборе диаметра. Клеммные колодки требуют, чтобы болтовые соединения были надежно затянуты.
Колпачки СИЗ в распределительной коробкеСкрутка в чистом виде не входит в перечень дозволенных ПУЭ соединенийСледует помнить, что согласно ПУЭ, скрутка проводов не допускается в чистом виде
Испытание соединений проводов в распределительных коробках
После выполнения всех соединений, оголенные участки проводников изолируют при помощи термоусадочных трубок, и укладывают провода в распределительной коробках. Сами коробки оставляют открытыми до проведения испытаний смонтированной электропроводки. Вначале на подключенные линии подается напряжение включением соответствующих автоматических выключателей.
Сами коробки оставляют открытыми до проведения испытаний смонтированной электропроводки. Вначале на подключенные линии подается напряжение включением соответствующих автоматических выключателей.
Если после включения нигде ничего не заискрило и автомат не выбило от короткого замыкания при ошибочном подсоединении проводов или некачественной изоляции соединений, проводят испытания электропроводки током нагрузки (прогрузка), которые производится при помощи подключения различных электроприборов на смонтированные линии. Рекомендуется нагрузить каждую линию максимально допустимым током.
Прогрузка должна продолжаться некоторое время (желательно несколько часов). За данный период возможные дефекты электромонтажа успеют себя проявить. Следует произвести визуальный осмотр соединений в распределительных коробках – признаки большой температуры будут видны по оплавлению изоляции или клеммников. Также важно отсутствие характерного запаха перегретой или горелой изоляции.
Также важно отсутствие характерного запаха перегретой или горелой изоляции.
После снятия напряжения следует на ощупь проверить все соединения – они не должны быть горячими. Если при прогрузке электропроводки максимальным расчетным током в течение нескольких часов не выявлено никаких замечаний по поводу работы соединений, то электромонтаж считается нормальным, распределительные коробки можно закрывать и вводить проводку в эксплуатацию.
Соединение проводов в распределительной коробке? СХЕМЫ
монтаж распределительной коробкиДля того чтобы обеспечить каждый уголок квартиры или офиса электричеством, не обойтись без распределительной коробки. Современные распределительные коробки очень разнообразны как по форме, так и по материалам,из которых они изготовлены. Они бывают накладной установки и внутренней.
Цель таких устройств заключается в том, чтобы развести провода в нужное направление к точкам потребления или размыкания — это могут быть выключатели, розетки или осветительные приборы. Развести провода в распределительной коробке профессиональному электрику не доставит хлопот, но вот для начинающего электромонтажника это может оказаться настоящей наукой. На сегодняшний день, для контактных соединений в распределительной коробке все чаще используют универсальные клеммы WAGO.
Соблюдаем расцветку проводов в распределительной коробке
расцветка проводовДля того чтобы справится с монтажем, нужно подключить воображение: провода это “трубы”, а электрический ток “вода”. По проводам фазы идет «подача воды”, а вот по нулевому проводнику течет “обратка” , тогда как защитный проводник служит на всякий аварийный случай: если где-то протечет “вода”, она непременно будет “слита” в землю. Очень удобно, что современные провода окрашены в различные цвета. Самая распространенная расцветка выглядит так: белый цвет – фаза (L), синий – нуль (N), желто-зеленый — заземление (РЕ).
Во время электромонтажа очень важно соблюдать последовательность в расцветке проводов, что очень облегчит их разведение в распределительной коробке. Прежде чем приступить к электромонтажным работам, нужно четко определиться с установкой точек освещения, розеток и выключателей, т.е. составить схему, чтобы установить распределительные коробки в удобных местах, и в будущем можно было осуществить профилактические и ремонтные работы.
Обязательно прочитайте подробные статьи про соединение проводов:
Порядок установки распределительной коробки
Соединение проводов в распределительной коробкеМногие делают ремонт, используя натяжные или подвесные потолки, поэтому электропроводку нужно прокладывать так, чтобы она не оказалась в зоне сверления отверстий когда будут монтироваться направляющие, а так же необходимо позаботится о том, чтобы распределительные коробки остались в зоне доступа. Если распределительная коробка осталась за подвесным потолком, можно установить маленький люк.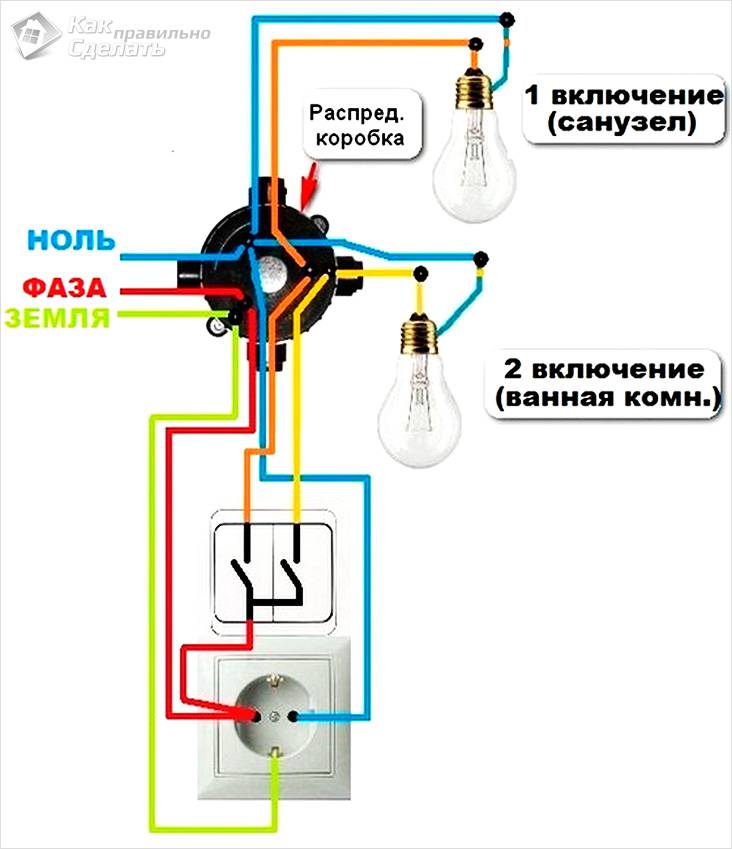
Последнее слово электротехники гласит так: от распределительного щита нужно прокладывать провод для освещения — 3х1,5, для розеток провод — 3х2,5. Поэтому, в распределительной коробке может оказаться достаточно большое количество проводов для соединения. Чтобы не запутаться, их необходимо маркировать.
Подключение проводов без заземления (РЕ)
Подключение проводов без заземления (РЕ)Подключение проводов с заземлением (РЕ)
Подключение проводов с заземлением (РЕ)Схема в распределительной коробке, двух светильников
Схема распределительной коробки на 2 светильникаСхема в распределительной коробке, двух светильников и группа розеток
Схема подключения на освещение и розеткиОцените качество статьи:
Распайка коробки
Расключение коробок
Расключение распределительной коробки (Распайка) производится опытным специалистом. Согласно ПЭУ, электрические соединения проводов должны выполняться только в распределительных коробках.
Обратившись в ООО «Эфас» вы можете быть уверенны в надежности, качестве, долговечности данной работы. Мы предоставляем гарантию на распайку распределительных коробок 5 лет. Очень часто можно встретить некачественное соединение проводов, из-за чего может произойти пожар, или просто пропадет напряжение в квартире, доме или на производстве. Помните, что сейчас «специалистов» очень много, но качественную распайку коробки выполнят не многие.
Распайка коробки
Расключение распределительных коробок осуществляется несколькими способами:
Пайка, опрессовка, сварка, использование клемм, Зажимов. Все эти способы можно использовать при расключении распределительных коробок. Самым надежным способом считается опрессовка и сварка, однако это занимает больше времени и требует специального оборудования и опыта.
Коммутация сети
Коммутация это процесс переключения в электрических цепях при замыкании или размыкании различных участков цепи. Каждый элемент электрической цепи (Розетка, выключатель, светильник) сводится в распределительную коробку. Потом в этой распределительной коробке необходимо соединить провода в нужном соотношении, чтобы выключатель был выключателем, а не розеткой и наоборот. Для этого нужно руководствоваться ПУЭ.
Каждый элемент электрической цепи (Розетка, выключатель, светильник) сводится в распределительную коробку. Потом в этой распределительной коробке необходимо соединить провода в нужном соотношении, чтобы выключатель был выключателем, а не розеткой и наоборот. Для этого нужно руководствоваться ПУЭ.
Распайка проводов в коробке
Основной шаблон действий при распайки проводов.
— Заводим кабеля в распределительную коробку (от розеток, выключателя, светильника, питающей линия)
— Обрезаем лишнюю длину кабеля (оставляем примерно 15-20 см)
— Снимаем верхний слой изоляции кабеля до основания коробки
— Снимаем изоляцию проводов нужной длины (примерно для клемм типа Wago 1,5-2 см, для скрутки-5 см)
— Производим соединение проводов согласно схемы
— Укладываем соединения в распределительную коробку и закрываем крышкой
Вот например стандартная схема
Схема коммутации
При расключении коробок руководствуйтесь электрической схемой. Как правило стандартный трехжильный кабель имеет следующую расцветку: белый, синий, желто-зеленый, где
Как правило стандартный трехжильный кабель имеет следующую расцветку: белый, синий, желто-зеленый, где
белый-фазный проводник и обозначается как «L»
синий-нулевой проводник и обозначается как «N»
желто-зеленый-заземляющий проводник обозначается как «PE»
Самый оптимальный способ будет при расключении коробок — это скрутка с использованием ЗАЖИМА СИЗ (самоизолирующий зажим) он будет поддерживать скрутку в надежном соединении при максимальных нагрузках.
Распайка электрических коробок
Расключение распределительных коробок требует особого внимания и должно производиться квалифицированным электриком. После выполнения электромонтажных работ рекомендуем Вам проверить на максимальной нагрузке данную группу (автомат на котором сидит питание коробки). В случаи некачественного расключения на высоких нагрузках вы можете обнаружить запах, искрение, моргание света и другие видимые дефекты из-за некачественного соединения проводов.
Комментарии и размещение обратных ссылок в настоящее время закрыты.
Расключение распределительной коробки — Всё о электрике
Как выполнить расключение коробки
В любом помещении как минимум имеется одна ответвительная коробка, именуемая распаечной. Иногда дополнительная может быть смонтирована на розетки – при условии их значительного удаления друг от друга. Расключение коробки необходимо при многих профилактических и ремонтных работах.
Вся проводка в жилых и бытовых зданиях прокладывается в полу или по стенам. Кабели и провода в гараже, загородном доме, квартире имеют выход из электрощита, где расположены счетчик, учитывающий потребление электроэнергии, предохранительные пробки или автоматы, и приходят к месту нахождения разветвительных и распаечных коробок. Здесь токопроводники скручиваются и соединяются способом использования специальных зажимов.
Далее проводка прокладывается уже от коробок к осветительным элементам, розеткам и выключателям. Встречаются и варианты соединения с другими коробками, подключенными для отдельных автоматов и групп потребителей.
Встречаются и варианты соединения с другими коробками, подключенными для отдельных автоматов и групп потребителей.
Обычно в жилое помещение старой застройки заводится 2 группы, а в новые квартиры – 3 группы. Отдельный ввод имеет жилье с установленными согласно проекту электроплитами. Токопроводники для каждой отдельно взятой группы проходят автономно, не имея точек соединения. При повреждении будет отключена только часть проводки, сидящая на своем автомате. Очень важно предусмотреть, чтобы в электрощите общая нагрузка между автоматами распределялась пропорционально.
Основное требование в мерах безопасности – обязательно выкручиваются пробки или отключается автомат для выполнения любых работ.
Последовательность выполнения монтажа коробки и ее расключения
Ознакомимся с пошаговой инструкцией правильного процесса установки:
- Выбиваем отверстия в коробке или используем уплотнители специального назначения в отверстиях для накладных для того, чтобы произвести внутреннюю установку.
 В этих местах заводим вовнутрь кабели или провода.
В этих местах заводим вовнутрь кабели или провода. - Два способа применяются для крепления коробки распределения. Первый – саморезами или парой дюбелей фиксируем ее на стене или потолке, как накладной элемент наружной проводки. Второй – Для скрытого исполнения заподлицо с поверхностью стены замазывается раствором распределительная коробка.
- Полностью снимается внешний изоляционный слой с кабелей на всем участке вплоть до входа в коробку. При этом следует соблюдать повышенную осторожность во избежание повреждения второй изоляции токопроводников. Желательно попробовать сделать это на пришедшем в негодность куске провода. При обнаружении поврежденных участков обмотайте их слоем изоленты.
- Еще один ответственный момент – при откусывании излишка жил или участка провода важно предусмотреть, чтобы остатка хватало для проведения скруток и укладки во внутреннюю часть коробки. Лично я предпочитаю делать запас длины в 15 см. В ситуации с большим количеством кабелей укорачиваю кабеля до 10-12 см.
- Определившись с нужной длиной, производим зачистку концов.
- В соответствии со схемой работы применяем электрозажимы для соединения проводов.
 В этих местах заводим вовнутрь кабели или провода.
В этих местах заводим вовнутрь кабели или провода.
Не так давно для получения соединений использовали метод скрутки и нанесения на это место слоя защитного покрытия. Подобная методика уступила место более усовершенствованному способу при помощи клеммников или СИЗ. Можно отыскать много материалов по описанию всех нюансов и особенностей данного процесса.
- Далее потребуется произвести размещение во внутренней части и разделение на отдельные группы пучков ноля или земли и фазы. В случае если токопроводники расположены достаточно свободно, применяется элементарный способ их проталкивания в требуемую зону коробки ручкой молотка.
- Выполнив все перечисленные процедуры, одеваем на место крышку.
Схема для выполнения расключения
Выполняется скручивание строго по цветам, обозначенным на рисунке, всех защитных проводников («Земля»), имеющих желто-зеленую окраску, и «Нолей» с изоляцией проводов синего исполнения. При двухпроводном варианте «Земля» отсутствует.
При двухпроводном варианте «Земля» отсутствует.
Немного по-другому происходит распаивание фазы с черным или красным цветом. Соединение вместе фазных проводников выполняется при варианте с выходом от коробки исключительно кабелей на розетки.
В ситуации с одноклавишным светильником потребуется один из фазных проводов, подведенных к выключателю, скрутить с L на выключателе – всеми фазными проводниками устройства. А затем фазный провод L на освещение, идущий на люстру, скрутить с проводником, отведенным от выключателя. В результате у нас получится 4 соединения.
Рассмотрим ситуацию с процессом расключения для трехпроводной электрической проводки двухклавишного устройства выключения. Потребуется прокладка четырехжильного кабеля (земля+ноль+1включение фаза+2 включение фаза) для люстры с двойным включением. В двухпроводном варианте достаточно трех жил, так как нет потребности в земле. Давайте посмотрим схему именно для последнего случая.
Как можно увидеть, в итоге получается четыре скрутки.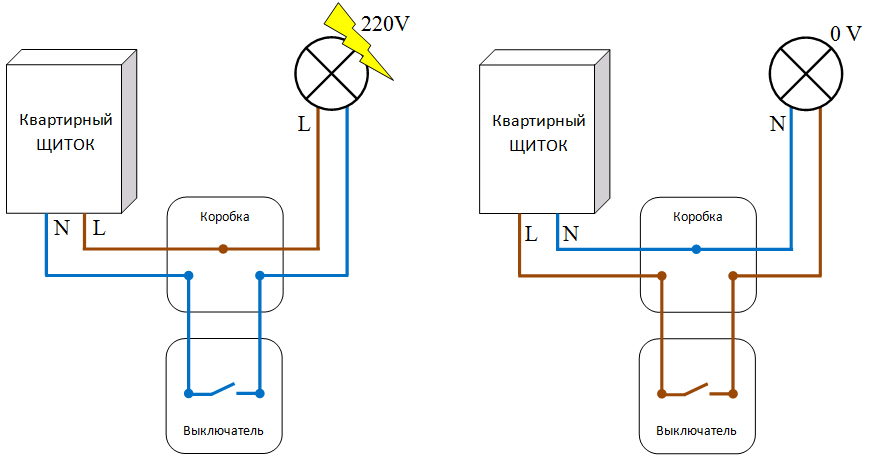 При этом мы не берем в расчет место соединения заземляющих проводников. Все без исключения ноли синего цвета должны быть соединены между собой.
При этом мы не берем в расчет место соединения заземляющих проводников. Все без исключения ноли синего цвета должны быть соединены между собой.
Также производится совместная скрутка фаз на розетках, на общий зажим выключателя двухклавишной конструкции с двумя проводниками, идущими на лампочки светильника независимо друг от друга, и питающего кабеля. Более подробно можно рассмотреть данный процесс в статье о подключении осветительных приборов.
Даже без большого опыта в электрических работах распаять коробку собственными силами не так уж и сложно. При обнаружении каких-либо неисправностей тщательно проверьте схему расключения.
Главное, что потребуется от вас – это четко придерживаться всех положений инструкции, проявлять осторожность и аккуратность в ходе работы. А также выполнять золотое правило профессиональных электриков – никогда не проявлять чрезмерной спешки, чтобы обезопасить себя от негативных моментов в процессе эксплуатации проводки.
Расключение распределительной коробки
Расключение распределительной коробки (Распайка) производится опытным специалистом. Согласно ПЭУ, электрические соединения проводов должны выполняться только в распределительных коробках.
Согласно ПЭУ, электрические соединения проводов должны выполняться только в распределительных коробках.
Обратившись в ООО «Эфас» вы можете быть уверенны в надежности, качестве, долговечности данной работы. Мы предоставляем гарантию на распайку распределительных коробок 5 лет . Очень часто можно встретить некачественное соединение проводов, из-за чего может произойти пожар, или просто пропадет напряжение в квартире, доме или на производстве. Помните, что сейчас «специалистов» очень много, но качественную распайку коробки выполнят не многие.
Распайка коробки
Расключение распределительных коробок осуществляется несколькими способами:
Пайка, опрессовка, сварка, использование клемм, Зажимов. Все эти способы можно использовать при расключении распределительных коробок. Самым надежным способом считается опрессовка и сварка, однако это занимает больше времени и требует специального оборудования и опыта.
Коммутация сети
Коммутация это процесс переключения в электрических цепях при замыкании или размыкании различных участков цепи. Каждый элемент электрической цепи (Розетка, выключатель, светильник) сводится в распределительную коробку. Потом в этой распределительной коробке необходимо соединить провода в нужном соотношении, чтобы выключатель был выключателем, а не розеткой и наоборот. Для этого нужно руководствоваться ПУЭ.
Распайка проводов в коробке
Основной шаблон действий при распайки проводов.
— Заводим кабеля в распределительную коробку (от розеток, выключателя, светильника, питающей линия)
— Обрезаем лишнюю длину кабеля (оставляем примерно 15-20 см)
— Снимаем верхний слой изоляции кабеля до основания коробки
— Снимаем изоляцию проводов нужной длины (примерно для клемм типа Wago 1,5-2 см, для скрутки-5 см)
— Производим соединение проводов согласно схемы
— Укладываем соединения в распределительную коробку и закрываем крышкой
Вот например стандартная схема
Схема коммутации
При расключении коробок руководствуйтесь электрической схемой. Как правило стандартный трехжильный кабель имеет следующую расцветку: белый, синий, желто-зеленый, где
Как правило стандартный трехжильный кабель имеет следующую расцветку: белый, синий, желто-зеленый, где
белый-фазный проводник и обозначается как «L»
синий-нулевой проводник и обозначается как «N»
желто-зеленый-заземляющий проводник обозначается как «PE»
Самый оптимальный способ будет при расключении коробок — это скрутка с использованием ЗАЖИМА СИЗ (самоизолирующий зажим) он будет поддерживать скрутку в надежном соединении при максимальных нагрузках.
Распайка электрических коробок
Расключение распределительных коробок требует особого внимания и должно производиться квалифицированным электриком. После выполнения электромонтажных работ рекомендуем Вам проверить на максимальной нагрузке данную группу (автомат на котором сидит питание коробки). В случаи некачественного расключения на высоких нагрузках вы можете обнаружить запах, искрение, моргание света и другие видимые дефекты из-за некачественного соединения проводов.
Способы соединения проводов в распределительной коробке
Способов соединения проводов в распределительных коробках, при монтаже электропроводки или просто удлинении электрических линий, достаточно много, но не все они разрешены, не все одинаково надежны. При этом, правила соединения однопроволочных проводов и многопроволочных, из-за их конструктивных отличий, различаются.
В первую очередь, выбирая способ соединения, нужно обратится к ПУЭ (Правила устройства электроустановок), основному документу, регламентирующему монтаж электропроводки.
Согласно пункту 2.1.21, из ПУЭ 7го издания:
Соединение, ответвление и оконцевание жил проводов и кабелей должны производиться при помощи опрессовки, сварки, пайки или сжимов (винтовых, болтовых и т. п.) в соответствии с действующими инструкциями, утвержденными в установленном порядке.
Единственное, что я бы добавил к этому – требования ГОСТ Р 50571.15-97, в котором не рекомендуется пайка проводов при соединении, поэтому, этот способ мы рассматривать не будем. Наша задача выбрать только самые надежные и удобные способы, пайка же, по этим параметрам, не подходит.
Наша задача выбрать только самые надежные и удобные способы, пайка же, по этим параметрам, не подходит.
Разрешенные способы соединения проводов
Опрессовка
Для соединения проводов методом опрессовки используются специальные гильзы, представляющие собой полые металлические трубки, в которые помещаются жилы соединяемых проводов, а затем, с помощью ручных пресс-клещей или механического пресса опрессовываются.
Принцип этого метода соединения, заключается в механическом сжатии всех соединяемых жил, что обеспечивает необходимый электрический контакт, а так же надежность и долговечность соединения.
Главным недостатком опрессовки проводов для соединения проводов в распределительных коробках, является необходимость использования гильз разного сечения и размеров, а также специализированного инструмента – пресс-клещей. Кроме того, в этом методе, достаточно сложно добавить к соединению еще проводники после опрессовки, а использование пресс-клещей, при монтаже проводки в квартире, не всегда удобно и возможно, так как для их работы, требуется определенное пространство, которое, к сожалению, в реальных условиях не всегда бывает.
Наиболее эффективно опрессовка показывает себя в соединении многопроволочных проводов, для однопроволочных (моножильных) она используется достаточно редко. Чаще всего ей пользуются в случае, когда у монтажника уже есть весь необходимый инструмент и опыт работы. Кроме того, опрессовка не требует электричества и может выполнятся в тех местах, где его нет.
Сварка
Принцип соединения проводов и кабелей сваркой, основан на сплавлении медных жил при воздействии высокой температуры от электрической дуги сварочного аппарата. При этом получается надежное соединение, при котором все жилы объединяются не межатомном уровне, соответственно и сопротивление у такого соединения рекордно низкое (не нагревается под нагрузкой).
В настоящий момент сварка считается наиболее надежным и качественным видом соединения однопроволочных медных жил проводов и кабелей, применяемых при монтаже проводки.
К главным недостаткам сварки, можно отнести необходимость наличия узкоспециализированного сварочного оборудования и навыка владения им у специалиста, выполняющего монтаж. Кроме того, производство сварочных работ требует свободного пространства в месте монтажа и самое главное, наличия электричества. Места соединений сварного соединения проводов в распределительной коробке также необходимо дополнительно изолировать.
Кроме того, производство сварочных работ требует свободного пространства в месте монтажа и самое главное, наличия электричества. Места соединений сварного соединения проводов в распределительной коробке также необходимо дополнительно изолировать.
Зажимной метод самый доступный из способов соединения проводов. Его принцип довольно прост, токопроводящие жилы кабелей или проводов, стягиваются, сжимаются, друг с другом, с помощью различного рода соединителей (винтовых, пружинных и т.д.). Самым ярким представителем этого способа соединения проводов являются клеммы.
Клеммы, для соединения жил при монтаже электропроводки, чаще всего бывают или винтовыми – где провода стягиваются в общей колодке винтами, или самозажимными – в которых жилы проводов зажимаются между подпружиненных пластин.
Винтовые клеммы, чаще всего используются для подключения электрооборудования, при соединении кабелей в распределительных коробках их не применяют. Одним из главных недостатков винтового соединения, является то, что со временем контакт ослабевает и винт необходимо затягивать. Если этого не делать место соединения начнет греться и в результате это может стать причиной возгорания либо нестабильной работы электросети.
Если этого не делать место соединения начнет греться и в результате это может стать причиной возгорания либо нестабильной работы электросети.
Самозажимные клеммы, на основе плоскопружинного зажима (крепеж под пружинящими пластинами), идеально подходят для соединения жил кабелей или проводов при монтаже электропроводки. Для того, чтобы соединить провода, достаточно поместить оголенные жилы в разъемы клемм, где они автоматически зафиксируются и будут связаны между собой через токопроводящий материал внутреннего механизма клеммы.
И хотя такое соединение получается не таким надежным как при сварке, используется оно при монтаже электропроводки очень часто. В первую очередь из-за простоты и быстроты монтажа. Достаточно только снимать изоляцию с жил кабелей и помещать их в клеммы.
Главным же недостатком такого способа соединения, является необходимость покупки качественных самозажимных клемм. Кроме того, довольно неоднозначно проявляют себя такие соединения во многих экстремальных ситуациях, которые, к сожалению, могут возникнуть в процессе эксплуатации электросети.
Опытные электромонтажники, стараются использовать самозажимные клеммы лишь на группы освещения, а соединения кабелей идущих, например, на розетки выполняют сваркой.
Если вы решили делать проводку в квартире своими руками, то соединение самозажимными клеммами для вас будет самым предпочтительным вариантом. Главное, это использовать клеммы, специально предназначенные для коммутации силовых цепей и рассчитанных на это. Еще одним плюсом является то, что такие соединения нет необходимости дополнительно изолировать, что так же очень сильно экономит время.
Еще есть клеммы с рычажковым зажимом, в них фиксация жилы происходить при закрытии рычага, а при его открытии она снова освобождается. Такую клемму можно использовать многократно, но они достаточно объемные и дорогие, поэтому используются при соединениях в распределительных коробках редко. Главное их преимущество над самозажимными клеммами – возможность соединять многопроволочные провода, без дополнительной подготовки жил.
Каким способом лучше всего соединять провода в распределительной коробке
Выше представленны все разрешенные методы, которыми можно соединять электрические провода в распределительной коробке, при монтаже проводки в квартире или доме. Каждый из них имеет свои особенности, сильные и слабые стороны. Очевидно, что выбор стоит делать между двумя:
1. Сварку проводов, однозначно нужно рассматривать в первую очередь, т.к. такой метод соединения проводов гарантирует максимальную надёжность всей проводки. Если вы хотите быть полностью уверенными в своей электросети, а как известно соединения самые узкие места любой электропроводки, я рекомендую взять сварочный аппарат в аренду, купить или собрать самому, чтобы иметь возможность сварить провода в коробках.
2. Соединение проводов самозажимными клеммами, подойдёт для тех, кто хочет сделать проводку быстро, при этом получив достаточно качественное соединение. Очень важно в таком случае, правильно рассчитать электропроводку и не перегружать её. Очень многие электромонтажники, не только самоучки, выбирают в последнее время именно клеммы, из-за простоты и скорости монтажа.
Очень многие электромонтажники, не только самоучки, выбирают в последнее время именно клеммы, из-за простоты и скорости монтажа.
Выполнять опрессовку, я бы рекомендовал лишь в случаях, когда у вас уже есть специализированное оборудование (пресс-клещи) и материал – гильзы.
А если вы знаете, еще удобные разрешенные способы соединения проводов при электромонтаже – оставляйте комментарии. Кроме того, напишите о вашем опыте, какому способу соединения и почему вы отдаёте предпочтение. Думаю, это будет полезно всем! Так же, как обычно, приветствуются любые комментарии по теме, вопросы, обсуждения!
{SOURCE}
Коробка испытательная переходная (КИП) — варианты подключения
Согласно принятым нормам, есть особая группа потребителей, которых нельзя отключать от питающей энергосистемы даже на непродолжительное время. Но что делать, когда для цепей учета необходимо произвести замену трехфазного счетчика или испытательная лаборатория, должна выполнить поверку при помощи эталонного устройства контроля?
При описанных выше условиях обратиться к первому разделу в своде правил установок электрооборудования.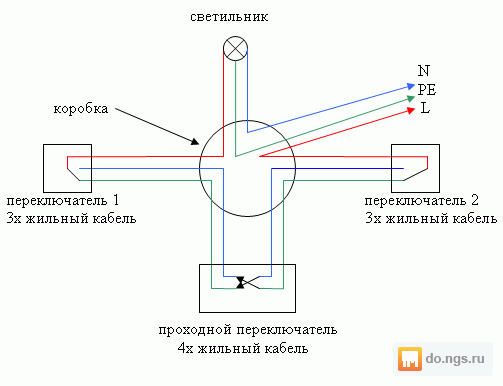 В нем указано, что для подключения счетчика с трансформатором тока (в тексте будет использована аббревиатура «ТТ»), должна устанавливаться переходная испытательная коробка, например, такая, как на рисунке 1.
В нем указано, что для подключения счетчика с трансформатором тока (в тексте будет использована аббревиатура «ТТ»), должна устанавливаться переходная испытательная коробка, например, такая, как на рисунке 1.
Назначение
Данное приспособление применяется, когда необходимо выполнить монтаж цепей учета на основе электросчетчиков с трансформаторным включением. Такое решение позволяет проделать работу, без обесточивания потребителей:
- подключать в щиток образцовое приспособление учета;
- производить шунтирование и отключение токовых цепей;
- выполнить расключение определенной фазы.
Первое действие выполняется, когда производится тестирование приспособлений контроля, остальные — при их замене.
Конструктивные особенности и основные характеристики
Рассмотрим, как устроен контактный бокс на примере КИ УЗ (см. рис.2)
Рисунок 2. Расположение контактов в ИКККонтакты с пометками 0, А, В и C используются для силовой цепи, а зажимы, имеющие номера с 1-го по 7-й служат для токового участка. Как выполняется включение КИП, будет рассказано в следующем разделе.
Как выполняется включение КИП, будет рассказано в следующем разделе.
Конструкция КИП представляет собой контактную группу, размещенную в пластиковой коробке из ударопрочного и негорючего поликарбоната. Размеры этой модели — 68х220х33 мм.
Параметры рабочего напряжения и тока – 380 В и 16 А. Изоляционные свойства материала позволяют выдерживать кратковременное превышение до 2000В и 25А. Для изготовления токоведущих частей используется латунь. Допускается ее замена оцинкованной сталью, но срок службы таких контактов становится короче. В связи с этим производители известных брендов отдают предпочтение латуни.
Остальные эксплуатационные характеристики:
- модуль может использоваться при температурном режиме от -40 С° до 60 С°;
- допустимая влажность – не более 98 %;
- для подключения используется провода с минимальным сечением 0,5 мм2 и максимальным – 4 мм2;
- данная модель выпускается со степенью защиты IP20;
- длительность срока эксплуатации — до 30 лет.

Некоторые модели (например, BTS или КИП-5/25) выпускаются с прозрачной крышкой (см. рис. 3). Учитывая, что приспособления данного типа подлежат обязательному опломбированию, такая конструктивная особенность имеет очевидные преимущества, поскольку позволяет контролировать состояние группы контактов.
Рисунок 3. Прозрачная крышка позволит вовремя заметить перегрев зажима при плохом контактеВариант подключения
На рисунке 4 показана наиболее распространенная схема подключения приспособления учета, при помощи КИП.
Рисунок 4. Типовое подключение трехфазного приспособления учетаОбозначения:
- T1, T2, T3 – трансформаторы тока;
- Сч1 – трехфазное приспособление учета;
- К1 – бокс, через который выполняется подключение приспособления контроля.
Особенности схемы:
На рисунке 4 показано, что три фазы и нулевой провод подключаются к соответствующим местам на боксе и идут от него, непосредственно, к приспособлению учета. Очень важный фактор в данном случае – чередование фаз, оно не должно быть нарушено.
Очень важный фактор в данном случае – чередование фаз, оно не должно быть нарушено.
При подключении трех ТТ к боксу используется тип соединения «звезда».
Перемычки следует установить также, как продемонстрировано на рисунке 4.
Как производится отключение и подключение приспособления учета или образцового устройства
Выполняя замену необходимо соблюдать очередность действий, начнем описание с процедуры отключения.
Как производить отключение?
Делается это в следующем порядке:
- необходимо зашунтировать токовую цепь, чтобы сделать это, следует вкрутить в обозначенные на рисунке 5 места винты с соответствующей резьбой (как правило, м4). С обратной стороны бокса находится заизолированная шина, винтовое соединение обеспечит надежный контакт с ней.
Рисунок 5. Места, куда необходимо вкрутить винты
- Отключаются перемычки, указанные на рисунке 6. При этом, не обязательно их полностью снимать. Достаточно ослабить винты «a» «b» и «c» и перемычки можно будет разомкнуть. Рисунок 6. Перемычки обведены красным овалом, винты, которые нужно ослабить – синими стрелками
- Размыкаются перемычки в цепи напряжения, их расположение показано на рисунке 7. Рисунок 7. Для отключения силовой части необходимо снять отмеченные красным овалом перемычки
- На завершающем этапе производится отключение от бокса приспособления учета.
 С обратной стороны бокса находится заизолированная шина, винтовое соединение обеспечит надежный контакт с ней.
Рисунок 5. Места, куда необходимо вкрутить винты
С обратной стороны бокса находится заизолированная шина, винтовое соединение обеспечит надежный контакт с ней.
Рисунок 5. Места, куда необходимо вкрутить винтыПодключение нового устройства учета.
После того, как выполнен полный демонтаж, можно приступать к процедуре установки, выполняется она в обратном порядке, а именно:
- Производится монтаж приспособления.
- Выполняется подключение к боксу.
- Производится осмотр бокса на предмет, установлен ли шунт, если нет, то вкручивает соответствующие винты (см. рис. 5).
- К коробке подключается обмотка ТТ.
- Устанавливаются в рабочее положение перемычки в токовой и силовой зонах бокса (рисунок 6 и рисунок 7).
- Снимается шунтирование.
 рис. 5).
рис. 5).Зачем необходимо шунтирование?
Считаем необходимым дать небольшое пояснение о необходимости замыкать выходную катушку ТТ. Это связано с характерными особенностями таких устройств, нельзя допустить работу ТТ на холостом ходу с разомкнутой вторичной обмоткой. Если данное условие не будет выполнено,- на ней наведется большая ЭДС, что может не только привести к межвитковому замыканию, а и представлять опасность для жизни или здоровья человека.
Подключение образцового приспособления.
Алгоритм действий в такой ситуации примет следующий вид:
- Необходимо замкнуть выходы ТТ.
- Снять токовые перемычки с бокса.
- Отключить силовую часть.
- Подключить к боксу образцовое приспособление.
- Включить силовую часть.
- Отключить замыкающую шину.
- После проведения замеров образцовое устройство отключается и включается штатное, как это сделать было описано выше.

Для проведения тестового замера совершенно не обязательно отключать приспособление контроля от бокса. Особенности конструкции позволяют выполнить подключение, не снимая тестируемое устройство. Для этого контрольное приспособление подключается к нижним контактным группам бокса, а токовые перемычки не устанавливаются на место. В результате, штатное приспособление учета останется на месте, но не будет подключено к ТТ.
Теоретически, можно и не отключать токовые перемычки, но тогда будет довольно велика вероятность влияния штатного устройства на показания образцового приспособления.
Что необходимо принимать во внимание при работе с КИП?
На подключенном испытательном боксе имеется напряжение, опасное для человеческой жизни. Поэтому, для работы с этим устройством необходимо иметь соответствующий уровень допуска (до 1000 вольт).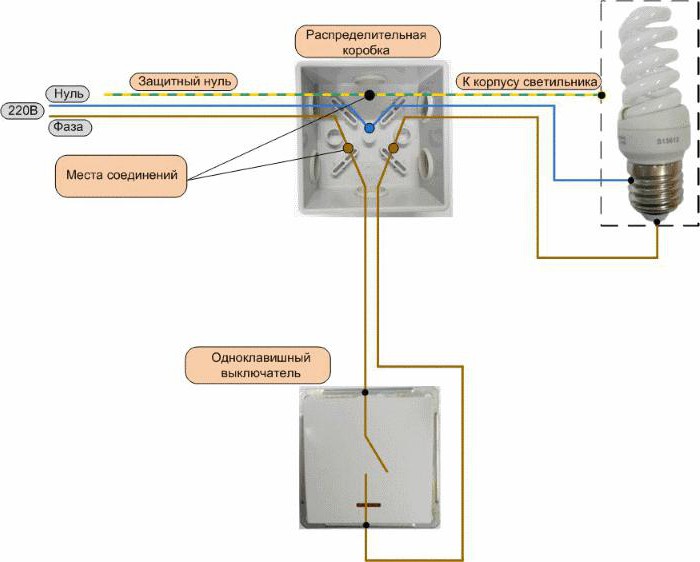
Поскольку данное приспособление подлежит обязательному опломбированию, то для манипуляций с ним могут быть допущены только лица, имеющие разрешение на проведение таких работ. Когда коммутация будет выполнена, бокс снова опечатывается.
Распределительные коробки. Виды и особенности установки
В правильном построении электрической проводки важную роль играют распределительные коробки. С помощью таких коробок осуществляется разводка линий питания электроэнергией по разным помещениям.
Изделие предназначено для обеспечения надежной изоляции соединительных узлов электропроводки с основным кабелем питания. Распределительные коробки конструктивно выполнены в виде пластикового корпуса со специальными отверстиями по бокам для прохода проводов, имеют закрывающуюся крышку.
Классификация
распределительные коробки классифицируют по методу установки:
- Для скрытой линии проводов. Коробка применяется в пустотах, проемах для организации места соединений проводов. Провода прокладываются в особых каналах, скрыты под штукатуркой, снаружи не видны. После установки и соединения проводов крышку закрывают и замазывают штукатуркой или шпаклевкой.
 Провода прокладываются в особых каналах, скрыты под штукатуркой, снаружи не видны. После установки и соединения проводов крышку закрывают и замазывают штукатуркой или шпаклевкой.
Провода прокладываются в особых каналах, скрыты под штукатуркой, снаружи не видны. После установки и соединения проводов крышку закрывают и замазывают штукатуркой или шпаклевкой.- Для наружного применения. Коробка закрепляется снаружи на поверхности стены. Там же располагают и кабель каналы для установки электропроводки. Этот вариант имеет место в офисной линии проводов, а также коммерческих организаций и промышленных предприятий. В частном строительстве наружная линия питания чаще предназначена для технических подсобных помещений. Наружный способ прокладки проводов удобен, так как можно быстро найти необходимое место, где соединены провода. Это является оптимальным решением для мест со значительным количеством потребителей энергии.
Данная классификация определяет только небольшую часть признаков распредкоробок, но является основной для широкого круга потребителей. Кроме этого вида классов, имеется разделение по назначению и степени безопасности распредкоробок. Этот способ деления актуален в помещениях спецназначения, дает максимальную возможность выбора изделий для создания безопасности, защитить электрооборудование от различных воздействий внутри зданий и помещений.
Этот способ деления актуален в помещениях спецназначения, дает максимальную возможность выбора изделий для создания безопасности, защитить электрооборудование от различных воздействий внутри зданий и помещений.
Классификация по виду защиты, особенностям и назначению:
- Распределительные коробки с обычной защитой. Применяются в производственных и жилых помещениях, служат для бытового применения, оснащены крышкой без уплотняющей прокладки, отверстия вывода проводов выполнены с уплотнением из резины.
- Пылезащитный вид. Служит для монтажа в пылесодержащих помещениях. Предохраняет соединение проводов от пагубного воздействия пыли, и как следствие, на функционирование системы обеспечения электроэнергией.
- Влагозащищенные коробки. Обеспечивают защиту соединений проводов от влаги и воды, с помощью резиновой прокладки на крышке, в помещениях с повышенной влажностью воздуха.
- Огнестойкого исполнения. Их использование актуально в помещениях с проведением взрывоопасных и огневых (сварочных) работ, с легко воспламеняемыми газами, жидкостями и веществами, с высокими требованиями безопасности. В огнестойких коробках провода монтируются с помощью специальных креплений, удерживаются специальными зажимами.
 Их использование актуально в помещениях с проведением взрывоопасных и огневых (сварочных) работ, с легко воспламеняемыми газами, жидкостями и веществами, с высокими требованиями безопасности. В огнестойких коробках провода монтируются с помощью специальных креплений, удерживаются специальными зажимами.
Их использование актуально в помещениях с проведением взрывоопасных и огневых (сварочных) работ, с легко воспламеняемыми газами, жидкостями и веществами, с высокими требованиями безопасности. В огнестойких коробках провода монтируются с помощью специальных креплений, удерживаются специальными зажимами.Коробки создают хорошую изоляцию для соединений электропроводки и от негативного действия внешних факторов, при этом имеют компактный корпус.
Многие распредкоробки состоят из пластмассового корпуса в виде прямоугольника или круга, разных типоразмеров, с отверстиями специального назначения для входа проводов внутрь коробки.
Распределительные коробки наружного исполнения выполнены с различными вариантами крепления их на поверхности стены, либо на других поверхностях с:
- Наружными установочными отверстиями.
- Внутренними крепежными отверстиями.
- Фиксацией на тросу. Имеют крепежные элементы фиксации на тросу.

Коробки для внутренней установки отличаются от наружных вариантов. Они устанавливаются внутри полых конструкций из гипсокартона, либо внутри стены. Крепление распредкоробок в стенах из бетона, кирпича или пенобетона производится шпаклевкой, раствором. Для надежности крепления на наружной стороне коробки выполнены удерживающие борозды.
Установка в гипсокартонных перегородках выполняется винтами в монтажные отверстия на дне коробки, либо специальными зажимами и элементами с распором конструкции.
Крышка, закрывающая корпус, фиксируется винтами или на защелках. Коробки внутреннего варианта имеют защелкивающуюся крышку, а наружный вариант выполнен с креплением крышки на винтах. Некоторые виды распредкоробок оснащены элементами для использования заземления, оснащены внутренней металлической планкой с крепежными винтами, отверстиями подключения кабеля и заземления.
Свойства и размерыДля монтажа элелектропроводки необходимо выбирать распредкоробку, которая бы удачно подходила для установки в определенном вами месте. Ее характеристика должна соответствовать требованиям и техусловиям.
Чаще всего к распределительным коробкам предъявляются следующие требования:
- Материал изготовления.
- Габаритные размеры, и размеры внутренней части.
Типоразмеры основных вариантов распредкоробок и пластмассы имеют много особенностей из-за технических условий изготовителя. Большинство вариантов типоразмеров круглой формы из пластмассы для наружного и внутреннего монтажа имеют:
- Диаметр 65 мм, глубина 40 мм, с классом защищенности IР55.
- 70 мм, глубиной – 50 мм.
- 80 мм, глубиной – 50 мм.
Такие детали оснащены четырьмя вводами для кабеля с заглушками из резины, уплотняющие вхождение кабеля. Крышка крепится за счет плотного облегания наружного диаметра коробки.
Пластмассовые коробки для проводов в виде прямоугольного корпуса выполняют со степенью защиты IР55, размеры сторон прямоугольника выполнены в интервале от 43 до 380 мм, с глубиной коробки от 35 до 120 мм.
Чаще всего для выпуска изделий такого вида, как распредкоробки, применяется пластик различного вида. Пластмасса имеет свои достоинства при изготовлении распредкоробок:
- Не проводит электрический ток.
- Невысокая стоимость.
- Простота технологического решения изготовления разных форм и размеров.
- Несложная возможность комплектования коробок вспомогательными элементами из резиновых прокладок, установочных планок, легкая установка крепежных винтов.
- Длительный срок службы коробки, обусловлен свойством пластмассы (стойкость к естественному разрушению).
При прокладке электропроводов на складах, мастерских и других помещениях, требующих не только хорошей фиксации проводов, но и защиту от негативных производственных факторов, появляется нужда в монтаже распредкоробок с повышенными свойствами в эксплуатации.
В этом случае оптимальным решением по выбору материала коробки становится металл.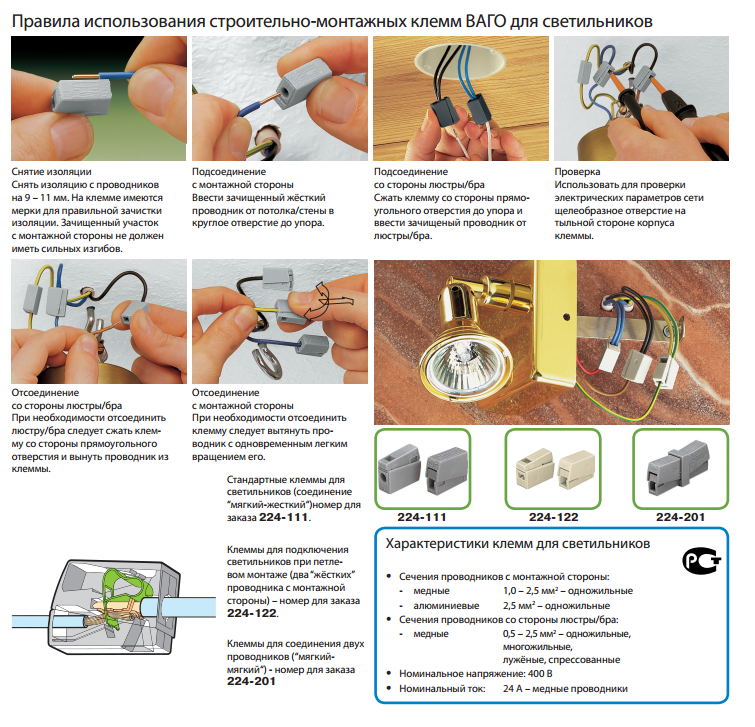 Коробки из металла изготавливаются чаще всего способом штамповки из листа стали, с дальнейшей грунтовкой и покраской. Крепление крышки создается применением винтового соединения. Конструкцию корпуса коробки делают герметичной при помощи уплотняющих резиновых прокладок в местах ввода электрокабеля, и прокладки из резины для крышки.
Коробки из металла изготавливаются чаще всего способом штамповки из листа стали, с дальнейшей грунтовкой и покраской. Крепление крышки создается применением винтового соединения. Конструкцию корпуса коробки делают герметичной при помощи уплотняющих резиновых прокладок в местах ввода электрокабеля, и прокладки из резины для крышки.
Распределительные коробки необходимы в местах, где разделяется центральный электрический кабель на отводящие линии потребителей. Согласно проекта устанавливаются определенные места (разветвления) монтажа распредкоробок при прокладке электропроводки. Чаще определяются места возле косяков в углу, или на расстоянии 30 см от потолка.
Порядок установки коробок для соединений проводов- В точке входа центрального кабеля в помещение монтируется распредкоробка, фиксируется на подготовленном месте.
- Провода отводов питания для потребителей различного назначения заводятся в корпус коробки на 5-7 см.
- Внутри корпуса коробки производится маркировка проводов.
- В случае необходимости делается чертеж схемы электропроводки на плане помещения, с указанием номеров проводов и особенностей прокладки.
- Производится скрутка проводов.
- Скрутки проводов чаще всего изолируют пластмассовыми колпачками (СИЗ) или WAGO.
- Скрутки помещаются внутрь корпуса коробки так, чтобы крышка могла закрыться.
- Закрывается крышка на корпусе.
- Производятся облицовочные работы стены, тем самым закрепляются распредкоробки шпаклевочным раствором или штукатуркой.
Установка соединений сети электропроводки в окончательную схему системы электроснабжения выполняется по подготовленной схеме. Нагрузка и толщина проводов рассчитываются в период проектирования.
Соединение проводов имеет свои особенности, поэтому нужно соблюдать некоторые правила:
- Концы провода очищают от изоляции на 2 см.
- Если кабели электропроводки оснащены двойной изоляцией провода, то наружный слой очищается на расстоянии 5 см.
- Соединение проводов выполняется по цветовому соответствию. Это намного облегчает трудоемкость монтажа проводки.
- С использованием плоскогубцев или другого инструмента для электрического монтажа проводки производят обжимку каждой скрутки — (СИЗ) или WAGO.
- В комплекте коробки могут поставляться специальные монтажные или клеммные колодки. При их наличии установка соединений проводов делается с применением таких элементов с винтовыми зажимами, не забывая о соответствии цветов проводов.
- После проведения , и окончания работы соединения сверяются со схемой электросети, узлы соединений укладываются в корпус коробки, закрывают крышку.

Похожие темы:
Блейк Микоски — ТОМС, процесс Хоффмана, сознательное разъединение и психоделики (# 446) — Блог автора Тима Феррисса
Иллюстрация через 99designsМастер в искусстве жизни не делает резких различий между своей работой и своей игрой; его труд и его досуг; его разум и его тело; его образование и его отдых.
— Л. П. Джекс
 Он вряд ли знает, что есть что. Он просто преследует свое видение совершенства во всем, что он делает, и оставляет других определять, работает он или играет.Самому себе, кажется, он всегда делает и то, и другое.
Он вряд ли знает, что есть что. Он просто преследует свое видение совершенства во всем, что он делает, и оставляет других определять, работает он или играет.Самому себе, кажется, он всегда делает и то, и другое.Блейк Микоски (@BlakeMycoskie) — серийный предприниматель, филантроп и автор бестселлеров, наиболее известный как основатель TOMS Shoes. Он также является автором идеи One for One®, бизнес-модели, которая помогает нуждающемуся человеку с каждым приобретенным продуктом. С момента основания TOMS Shoes предоставила детям по всему миру почти 96 миллионов пар обуви.
В 2014 году, после продажи половины компании Bain Capital, Блейк ушел с поста генерального директора TOMS.Используя половину своих доходов, он основал Фонд социальных предпринимателей, чтобы помочь молодым стартапам с ключевыми социальными миссиями начать работу с столь необходимым финансированием. С тех пор он инвестировал более чем в 25 социальных предприятий.
Весной 2020 года Блейк стал соучредителем своей новейшей компании Madefor , которая предлагает 10-месячную программу, в которой применяются принципы современной нейробиологии, психологии и физиологии, чтобы улучшить ваш мозг и тело. Madefor, созданный совместно с учеными из Стэнфорда, Гарварда и других ведущих университетов, помогает людям усваивать и поддерживать положительные привычки и практики, оказывающие наибольшее влияние на их жизнь.Слушатели этого подкаста могут использовать код TIM , чтобы получить 20% скидку на полные и ежемесячные планы.
Как автор бестселлеров New York Times книги « Start Something That Matters » 2011 года, Блейк предложил свою собственную историю вдохновения и силы включения пожертвований в бизнес. Блейк также недавно расширил свои филантропические усилия, включив в него финансирование Центра психоделических исследований и исследований сознания при Джонсе Хопкинсе, что сделало его первым подобным исследовательским центром в США и крупнейшим в своем роде в мире.
Слушайте выпуск в Apple Podcasts, Spotify, Overcast, Stitcher, Castbox, Google Podcasts или на своей любимой платформе для подкастов.
Предоставлено вам LMNT, Ascent Protein и Readwise. Подробнее обо всех трех ниже.
Стенограмму этого эпизода можно найти здесь. Стенограммы всех серий можно найти здесь.
Этот выпуск предоставлен вам LMNT! Что такое LMNT? Это вкусных напитков с электролитами без сахара. Я запасся коробками и коробками с этим веществом и обычно использую его 1-2 раза в день. LMNT разработан, чтобы помочь любому, у кого есть потребности в электролитах, и идеально подходит для людей, соблюдающих кето, низкоуглеводную или палеодиету. Если вы придерживаетесь низкоуглеводной диеты или голодания, электролиты играют ключевую роль в облегчении голода, судорог, головных болей, усталости и головокружения.
LMNT подготовил для вас особое предложение, мои дорогие слушатели. Они создали Клуб Тима: Просто зайдите на DrinkLMNT.com / tim, выберите «Подписаться и сэкономить» и используйте промокод TIMSCLUB, чтобы получить коробку LMNT с 30 счетами всего за 36 долларов. Это будет действовать в течение всего срока действия подписки, и вы можете приостановить ее в любое время.
Они создали Клуб Тима: Просто зайдите на DrinkLMNT.com / tim, выберите «Подписаться и сэкономить» и используйте промокод TIMSCLUB, чтобы получить коробку LMNT с 30 счетами всего за 36 долларов. Это будет действовать в течение всего срока действия подписки, и вы можете приостановить ее в любое время.
Этот эпизод также представлен вам Ascent Protein! Я использую Ascent в качестве основного протеинового порошка в течение последних трех лет. . Он чистый, эффективный и легкий для моего желудка. Многие протеиновые порошки содержат избыток сахара, сомнительные искусственные подсластители или низкосортный белок.Исключением является Ascent Protein: он не содержит искусственных ингредиентов, искусственных подсластителей и сахара. Все продукты Ascent сертифицированы Informed-Sport как не содержащие запрещенных веществ, а одна небольшая мерная ложка содержит 25 г белка и больше восстанавливающих мышцы BCAA, чем эквивалентная порция курицы, говядины, яиц, лосося или растительных белков.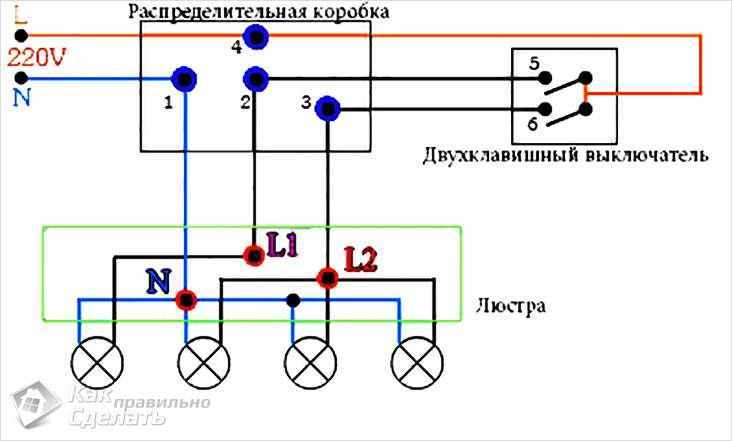 Посетите AscentProtein.com/tim и используйте код TIMASCENT для получения 20% скидки. Код скидки также будет работать на Amazon.com, если вы покупаете Ascent Protein.
Посетите AscentProtein.com/tim и используйте код TIMASCENT для получения 20% скидки. Код скидки также будет работать на Amazon.com, если вы покупаете Ascent Protein.
Этот эпизод также предлагает вам Readwise ! Readwise — это приложение, которое поможет вам запомнить значительно больше из того, что вы читаете. Как часто вы читаете книгу или статью, а затем через пару недель оглядываетесь назад и понимаете, что почти ничего из них не помните? Readwise решает эту проблему, интегрируясь с Kindle, Pocket, iBooks, Instapaper и т. Д., Чтобы отправлять вам ежедневный дайджест всех ваших основных моментов.В среднем их пользователи сообщают, что запоминают на 84% больше того, что они прочитали и подчеркнули.
Это помогает вам выработать увлекательную ежедневную привычку просматривать — и фактически использовать — сотни ярких моментов, которые просто сидят на ваших устройствах для чтения, собирают киберпыль и ничего не делают для вас. Эта простая привычка ежедневно пересматривать свои основные моменты может значительно улучшить объем информации, который вы сохраняете благодаря научно доказанному процессу интервального повторения. Зарегистрируйтесь по телефону по прочтениюio / tim за двухмесячная бесплатная пробная версия предлагается исключительно Tim Ferriss Show слушателям.
Эта простая привычка ежедневно пересматривать свои основные моменты может значительно улучшить объем информации, который вы сохраняете благодаря научно доказанному процессу интервального повторения. Зарегистрируйтесь по телефону по прочтениюio / tim за двухмесячная бесплатная пробная версия предлагается исключительно Tim Ferriss Show слушателям.
Какая ваша любимая цитата или урок из этого эпизода? Пожалуйста, дайте мне знать в комментариях.
ПРОКРУТИТЕ НИЖЕ ССЫЛКИ И ПОКАЗАТЬ ЗАМЕТКИ…
Хотите послушать Блейка в последний раз в сериале? Прислушайтесь к нашему разговору, в котором мы обсуждаем первые предпринимательские начинания, советы начинающим производителям, важность ведения журнала, органический рост по сравнению с привлечением венчурного капитала и многое другое.
ИЗБРАННЫЕ ССЫЛКИ ИЗ ЭПИЗОДА
- Связаться с Блейк Микоски :
Веб-сайт | Twitter | Facebook | Instagram
ПОКАЗАТЬ ЗАМЕТКИ
- Что побудило Блейка с ранних лет начать свою жизнь серийного предпринимателя? [06:39]
- Благодаря мечте стать профессиональным теннисистом, что сделало Блейка достаточно хорошим, чтобы стать соперником? [09:27]
- После многих лет работы на полную мощность над несколькими проектами, почему Блейк решил взять творческий отпуск в 2012 году — и что это повлекло за собой? Чувствует ли он, что переживал, ценно? [11:47]
- Что такое Процесс Хоффмана и почему Блейк решил участвовать в нем? [15:34]
- Как я подавляю реакцию кортизола перед сном, чтобы я действительно мог заснуть. [21:00]
- Какие разрушительные паттерны в его собственной жизни «Процесс Хоффмана» позволил Блейку выявить и обработать? [22:12]
- Блейк рекомендует книги и ресурсы, которые могут помочь людям достичь некоторых из того, что они могли бы пройти через Процесс Хоффмана. [26:22]
- Блейк разделяет сложный этап, который он пережил, проходя процесс Хоффмана. [29:28]
- Помимо Процесса Хоффмана, какие еще процессы, которые Блейк испробовал, оказали длительное влияние? [32:55]
- Как Блейк активно поддерживает научные исследования, связанные с психоделическими препаратами, что он извлек из собственного опыта в медицине растений? [36:08]
- Был ли у Блейка трудный или отрицательный опыт в медицине растений? [40:59]
- Объявление для всех, кто рассматривает возможность употребления психоделиков по какой-либо причине.[46:52]
- Что такое сознательное разъединение, как Блейк познакомился с ним и как это помогло ему лично — особенно в эпоху COVID-19? [50:45]
- Что еще заставило Блейка сфокусироваться на сознательном разъединении и позволило ему увидеть — возможно, впервые? [56:27]
- Когда во время отношений Блейк считает, что самое подходящее время для кого-то прочитать « Сознательное разъединение » Кэтрин Вудворд Томас? Могут ли они что-то сделать, прежде чем эти отношения явно перейдут на опасную территорию? Какие еще книги можно было бы прочитать сейчас? [1:00:31]
- Чего Блейк надеется достичь с помощью Madefor, своего нового бизнеса, который помогает людям учиться и поддерживать положительные привычки и практики, которые оказывают наибольшее влияние на их жизнь? Как это началось? [1:03:25]
- Каким образом Madefor выглядит с точки зрения потребителя? Какие привычки и практики представлены? [1:07:32]
- Прощальные мысли. [1:19:27]
 [21:00]
[21:00] [1:19:27]
[1:19:27]УКАЗАННЫЕ ЧЕЛОВЕКИ
Сопутствующие и рекомендуемыеШоу Тима Ферриса — один из самых популярных подкастов в мире, его скачали более 600 миллионов раз. Он трижды был выбран в категории «Лучшие подкасты Apple», часто является подкастом с интервью №1 среди всех подкастов Apple и неоднократно занимал первое место из более чем 400 000 подкастов. Чтобы послушать любой из прошлых выпусков бесплатно, посмотрите на этой странице .
IJMS | Бесплатный полнотекстовый | Моделирование молекулярной динамики митохондриального разобщающего белка 2
1. Введение
Разобщающий белок 2 (UCP2) принадлежит к митохондриальному суперсемейству SLC25 переносчиков анионов. Он был вовлечен в патогенез множества физиологических и патологических процессов, таких как диабет, ишемия, метаболические нарушения, (нейро) воспаление, рак и старение. Основываясь на своей функции транспорта протонов, UCP2 впервые предположили, что он действует как мягкий разобщитель для снижения окислительного стресса [1,2,3]. Позже было показано, что он может транспортировать метаболиты C4 из митохондрий [4], облегчая цикл трикарбоновых кислот (TCA). Недавно предложенная двойная транспортная функция для UCP2 (протон и субстрат) увеличивает сходство UCP2 с ANT (также сокращенно AAC в литературе), который транспортирует протоны [5,6,7], в дополнение к обмену ATP / ADP. То, как UCP2 контролирует транспорт протонов через митохондриальные мембраны, все еще не понятно. К настоящему времени установлено, что длинноцепочечные жирные кислоты (ЖК) являются неотъемлемой частью механизма и имеют решающее значение для переноса протонов [8,9,10].В настоящее время существует несколько механистических моделей, объясняющих механизм переноса протона. В первой, так называемой модели «циклирования ФА», ЖК действуют как протонофоры. Из-за избытка протонов в митохондриальном межмембранном пространстве, карбоксильные анионы FA легко протонируются, и они могут очень быстро перемещаться через мембрану в нейтральной форме к матрице [11,12,13], где впоследствии высвобождается протон.
Позже было показано, что он может транспортировать метаболиты C4 из митохондрий [4], облегчая цикл трикарбоновых кислот (TCA). Недавно предложенная двойная транспортная функция для UCP2 (протон и субстрат) увеличивает сходство UCP2 с ANT (также сокращенно AAC в литературе), который транспортирует протоны [5,6,7], в дополнение к обмену ATP / ADP. То, как UCP2 контролирует транспорт протонов через митохондриальные мембраны, все еще не понятно. К настоящему времени установлено, что длинноцепочечные жирные кислоты (ЖК) являются неотъемлемой частью механизма и имеют решающее значение для переноса протонов [8,9,10].В настоящее время существует несколько механистических моделей, объясняющих механизм переноса протона. В первой, так называемой модели «циклирования ФА», ЖК действуют как протонофоры. Из-за избытка протонов в митохондриальном межмембранном пространстве, карбоксильные анионы FA легко протонируются, и они могут очень быстро перемещаться через мембрану в нейтральной форме к матрице [11,12,13], где впоследствии высвобождается протон. После этого UCP2 способствует очень медленному переносу отрицательно заряженной жирной кислоты по еще неизвестному механизму обратно в межмембранное пространство, и цикл начинается снова [1,8,14].Зависимость скорости транспорта H + от насыщения FA, длины цепи FA [9] и текучести мембраны [15] указывает на то, что транспорт FA — , вероятно, происходит на границе раздела белок-липид. Вторая группа моделей не работает. включают триггеры FA. Вместо этого он предполагает, что карбоксильные группы отрицательно заряженных аминокислот UCP могут принимать протон от FA и транспортировать его по гипотетическому каналу в UCP (модель «буферизации протонов FA») [16,17]. Альтернативно, анион FA связывается в полости внутри UCP (модель «FA шаттл»).После связывания протона с анионом FA происходит конформационное изменение, которое перемещает FA вместе с протоном, который впоследствии высвобождается в матрице митохондрий, и цикл повторяется [6,18]. В настоящее время существует консенсус относительно точного механизма того, как До работ UCP2 еще далеко, в основном из-за нехватки надежной структурной информации. Потенциальный прорыв в исследовании UCP2 произошел в 2011 г., когда была опубликована структура ЯМР UCP2 [19]. Теоретически структура должна была служить идеальной отправной точкой для всех потенциальных молекулярных симуляций и подробных структурных и механистических анализов.К сожалению, оказалось, что структура UCP2, выделенная из обычно используемого детергента додецилфосфохолина (DPC), не имеет функционального значения [20]. Более того, в настоящее время точно установлено, что детергенты на основе алкилфосфохолина дестабилизируют и денатурируют альфа-спиральные мембранные белки, что приводит к искажению вторичной структуры белка. Это поднимает важные вопросы о пригодности детергентов на основе алкилфосфохолина в качестве экстракционной среды для определения структуры мембранных белков методом ЯМР в растворе.В настоящее время все еще ведутся оживленные дискуссии о том, является ли нарушение структуры белка этими типами детергентов препятствием для дальнейшего понимания функции белка [21,22,23,24], или его все еще можно использовать для улавливания наиболее важных функциональные аспекты [25,26,27].
После этого UCP2 способствует очень медленному переносу отрицательно заряженной жирной кислоты по еще неизвестному механизму обратно в межмембранное пространство, и цикл начинается снова [1,8,14].Зависимость скорости транспорта H + от насыщения FA, длины цепи FA [9] и текучести мембраны [15] указывает на то, что транспорт FA — , вероятно, происходит на границе раздела белок-липид. Вторая группа моделей не работает. включают триггеры FA. Вместо этого он предполагает, что карбоксильные группы отрицательно заряженных аминокислот UCP могут принимать протон от FA и транспортировать его по гипотетическому каналу в UCP (модель «буферизации протонов FA») [16,17]. Альтернативно, анион FA связывается в полости внутри UCP (модель «FA шаттл»).После связывания протона с анионом FA происходит конформационное изменение, которое перемещает FA вместе с протоном, который впоследствии высвобождается в матрице митохондрий, и цикл повторяется [6,18]. В настоящее время существует консенсус относительно точного механизма того, как До работ UCP2 еще далеко, в основном из-за нехватки надежной структурной информации. Потенциальный прорыв в исследовании UCP2 произошел в 2011 г., когда была опубликована структура ЯМР UCP2 [19]. Теоретически структура должна была служить идеальной отправной точкой для всех потенциальных молекулярных симуляций и подробных структурных и механистических анализов.К сожалению, оказалось, что структура UCP2, выделенная из обычно используемого детергента додецилфосфохолина (DPC), не имеет функционального значения [20]. Более того, в настоящее время точно установлено, что детергенты на основе алкилфосфохолина дестабилизируют и денатурируют альфа-спиральные мембранные белки, что приводит к искажению вторичной структуры белка. Это поднимает важные вопросы о пригодности детергентов на основе алкилфосфохолина в качестве экстракционной среды для определения структуры мембранных белков методом ЯМР в растворе.В настоящее время все еще ведутся оживленные дискуссии о том, является ли нарушение структуры белка этими типами детергентов препятствием для дальнейшего понимания функции белка [21,22,23,24], или его все еще можно использовать для улавливания наиболее важных функциональные аспекты [25,26,27]. Несмотря на недавние разработки в области определения структуры мембранных белков, такие как солюбилизация мембранных белков без использования детергентов с использованием липидных частиц стирола и малеиновой кислоты [28,29] и крио-ЭМ, сохраняющих целостность липидной среды [30,31], работа с небольшими митохондриями переносчики и, в частности, разобщающие белки, остаются сложной задачей из-за их размера и низкого содержания в митохондриальных мембранах.Моделирование молекулярной динамики (МД) является привлекательным дополнительным вариантом для изучения мембранных белков при условии, что время отбора проб достаточно велико для адекватного анализа их динамики в мембранах [28,29,30]. Мембранные белки кодируются ca. 30% генома человека и их общее количество, по прогнозам, будет значительно выше [31]. Сегодня определено только около 1000 уникальных структур мембранных белков [32], что составляет небольшую часть от общего числа мембранных белков, обнаруженных у человека.Однако, поскольку все большее количество структур мембранных белков определяется методом ЯМР в растворе с использованием спорных алкилфосфонатов в качестве экстракционной среды [33], длительное моделирование МД (в микросекундном временном диапазоне) в сочетании с моделированием гомологии [34,35] часто представляет собой единственная доступная опция для изучения структуры и динамики мембранных белков. В настоящее время МД-моделирование мономерных белков UCP, о которых сообщается в литературе, в основном основано на доступной структуре ЯМР UCP2 [20,36].МД-моделирование белка UCP2, проведенное Zoonens и соавторами, показало, что детергент DPC индуцировал большие структурные деформации спиралей белка UCP2, которые, в свою очередь, создавали большой водный канал, облегчая тем самым постоянную утечку воды через белок [20]. Поскольку протонная проводимость недостижима в этих условиях, его экспериментальные измерения дополнительно подтвердили, что белок UCP2, экстрагированный с помощью детергента DPC, не является функционально значимым. Напротив, близкородственный белок UCP1, экстрагированный с использованием DPC, и структурно отличающийся детергент TX-100 оставались физиологически активными [20].Интересно, что недавнее исследование, показывающее олигомеризацию мономеров UCP2, не описало различий между структурами модели ЯМР и гомологии [37]. Мотивированное отсутствием соответствующих МД-моделирования мономерных структур, которые помогли бы расшифровать функцию белка UCP2 в мембранах, мы обратились к моделированию гомологии с использованием структуры митохондриального носителя ADP / ATP (ANT, код PDB 1OKC) [38] в качестве шаблона для моделирования UCP2 MD. ANT, член семейства митохондриальных белков-переносчиков SLC25 [39], также обнаруживается во внутренних митохондриальных мембранах.Его основная функция — обмен АДФ на АТФ через митохондриальные мембраны [40,41,42]. Однако сообщалось, что ANT также работает как переносчик протонов, подобно белкам UCP, по механизму, еще не определенному на молекулярном уровне [5,6,7]. Принимая во внимание идентичность последовательностей между ANT и UCP2, равную 24% [43], сходство в общей форме, содержащей шесть мембранных доменов, а также совместное использование функции транспорта протонов в митохондриях, мы выбрали структуру 1OKC в качестве стартовой матрицы для гомологии. моделирование и последующие микросекундные МД моделирования.Наконец, мы сравнили результаты моделирования с данными, полученными на модельных мембранах, восстановленных с помощью UCP2, чтобы подтвердить нашу модель MD.
Несмотря на недавние разработки в области определения структуры мембранных белков, такие как солюбилизация мембранных белков без использования детергентов с использованием липидных частиц стирола и малеиновой кислоты [28,29] и крио-ЭМ, сохраняющих целостность липидной среды [30,31], работа с небольшими митохондриями переносчики и, в частности, разобщающие белки, остаются сложной задачей из-за их размера и низкого содержания в митохондриальных мембранах.Моделирование молекулярной динамики (МД) является привлекательным дополнительным вариантом для изучения мембранных белков при условии, что время отбора проб достаточно велико для адекватного анализа их динамики в мембранах [28,29,30]. Мембранные белки кодируются ca. 30% генома человека и их общее количество, по прогнозам, будет значительно выше [31]. Сегодня определено только около 1000 уникальных структур мембранных белков [32], что составляет небольшую часть от общего числа мембранных белков, обнаруженных у человека.Однако, поскольку все большее количество структур мембранных белков определяется методом ЯМР в растворе с использованием спорных алкилфосфонатов в качестве экстракционной среды [33], длительное моделирование МД (в микросекундном временном диапазоне) в сочетании с моделированием гомологии [34,35] часто представляет собой единственная доступная опция для изучения структуры и динамики мембранных белков. В настоящее время МД-моделирование мономерных белков UCP, о которых сообщается в литературе, в основном основано на доступной структуре ЯМР UCP2 [20,36].МД-моделирование белка UCP2, проведенное Zoonens и соавторами, показало, что детергент DPC индуцировал большие структурные деформации спиралей белка UCP2, которые, в свою очередь, создавали большой водный канал, облегчая тем самым постоянную утечку воды через белок [20]. Поскольку протонная проводимость недостижима в этих условиях, его экспериментальные измерения дополнительно подтвердили, что белок UCP2, экстрагированный с помощью детергента DPC, не является функционально значимым. Напротив, близкородственный белок UCP1, экстрагированный с использованием DPC, и структурно отличающийся детергент TX-100 оставались физиологически активными [20].Интересно, что недавнее исследование, показывающее олигомеризацию мономеров UCP2, не описало различий между структурами модели ЯМР и гомологии [37]. Мотивированное отсутствием соответствующих МД-моделирования мономерных структур, которые помогли бы расшифровать функцию белка UCP2 в мембранах, мы обратились к моделированию гомологии с использованием структуры митохондриального носителя ADP / ATP (ANT, код PDB 1OKC) [38] в качестве шаблона для моделирования UCP2 MD. ANT, член семейства митохондриальных белков-переносчиков SLC25 [39], также обнаруживается во внутренних митохондриальных мембранах.Его основная функция — обмен АДФ на АТФ через митохондриальные мембраны [40,41,42]. Однако сообщалось, что ANT также работает как переносчик протонов, подобно белкам UCP, по механизму, еще не определенному на молекулярном уровне [5,6,7]. Принимая во внимание идентичность последовательностей между ANT и UCP2, равную 24% [43], сходство в общей форме, содержащей шесть мембранных доменов, а также совместное использование функции транспорта протонов в митохондриях, мы выбрали структуру 1OKC в качестве стартовой матрицы для гомологии. моделирование и последующие микросекундные МД моделирования.Наконец, мы сравнили результаты моделирования с данными, полученными на модельных мембранах, восстановленных с помощью UCP2, чтобы подтвердить нашу модель MD.3. Обсуждение
Отсутствие соответствующей структуры UCP мешает пониманию транспортных механизмов и биологических функций UCP2. Чтобы найти возможное лекарство от текущей ситуации, мы обратились к моделированию гомологии с использованием кристаллографической структуры ANT и экспериментальной структуры ЯМР UCP2 в качестве стартовых структур для длинных микросекундных МД-моделирования белков UCP2 (UCP2 h и UCP2 NMR ).С помощью микросекундного МД-моделирования мы показали, что структура UCP2 h практически водонепроницаема (P f = 2,0 × 10 −16 см 3 с −1 ) с коэффициентом осмотической проницаемости воды на три порядка ниже. по величине, чем соответствующая структура UCP2 ЯМР после 2 мкс времени моделирования МД (P f = 1,3 × 10 −13 см 3 с −1 ). Это является следствием образования сети солевых мостиков на матричной стороне белка в UCP2 h , образованной остатками аргинина / лизина и аспартата / глутамата, а также сохранения тройной симметрии между отрицательно заряженными остатками на стороне матрицы. (Мотив EG), похожий на водонепроницаемые белки ANT [44,56,57].Напротив, стабильное образование идентичных солевых мостиков и мотива EG на матричной стороне структуры UCP2 ЯМР не наблюдалось, что позволяет воде почти свободно диффундировать через белок. Наконец, мы показали, что АТФ связывается со всеми тремя остатками аргинина в полости UCP2 только в случае структуры UCP2 h , что согласуется с предыдущими экспериментальными наблюдениями [53,54]. Здесь следует упомянуть, что моделирование белков UCP2 с помощью МД не полностью сходится (что практически невозможно при нынешних вычислительных мощностях) и что структура UCP2 h , возможно, не является глобальным минимумом.Тем не менее, мы считаем, что представленная структура UCP2 h актуальна для будущих исследований, поскольку она хорошо соответствует имеющимся экспериментальным данным и что ее поведение достаточно хорошо описано. Описанная структура UCP2 h может быть легко перенесена на мембраны, образованные DOPC. , ДОФЭ и кардиолипин, которые в таком случае более реалистично описывают липидный состав внутренних митохондриальных мембран и наших модельных мембран. Однако на данном этапе мы считаем, что дальнейшее усложнение модельной системы не является необходимым, как мы показали ранее в МД-моделировании белков ANT в различных, более сложных условиях [44].Из-за высокой сложности систем мембранный белок / липидный бислой и необходимости чрезвычайно длительного времени моделирования наиболее вероятны ошибки, возникающие из-за неполного покрытия всего фазового пространства в гетерогенных системах, даже в микросекундных временных масштабах и относительно небольших бислоев.Чтобы проверить достоверность результатов МД моделирования в отношении функциональности UCP2, мы сравнили связывание активатора UCP2 AA с обеими моделями, UCP2 h и UCP2 NMR . Кроме того, мы измерили активацию рекомбинантного UCP2 из-за связывания AA.Большая проводимость мембраны UCP2 в присутствии AA по сравнению с UCP2-R60S, в котором предполагаемый сайт связывания AA — был модифицирован, полностью согласуется с результатами моделирования MD. Это убедительно показывает, что предложенная область связывания AA — вокруг R60 доступна только в структуре белка UCP2 h и что в UCP2-R60S предотвращается связывание AA — . Это показывает, что структура UCP2 h правильно предсказывает связывание AA — и чувствительна к селективным мутациям, таким как R60S.Напротив, аналогичное связывание FA со структурой UCP2 ЯМР , как экспериментальное, так и после 2 мкс времени моделирования МД, не является оптимальным, что дополнительно ставит под сомнение стабильность и физиологическое значение экспериментальной структуры ЯМР.
Предлагаемая MD модель UCP2 h может в значительной степени способствовать исследованию механизма транспорта протонов, опосредованного UCP2, в целом и сайта связывания белков для FA в частности. В настоящее время предложены два основных механизма для транспорта протонов, опосредованного UCP1 (см. Введение).Ключевое различие между гипотезами «цикличности FA» и «челноком FA» заключается в локализации сайта связывания для активирующей FA. Модель «FA шаттл» утверждает, что FA — связывается внутри поры с цитозольной стороны UCP1 и переносит протоны путем челночного перемещения от цитозольной к матричной стороне митохондрий [58]. Напротив, гипотеза «цикла FA» предполагает, что FA — связывается с белком со стороны матрикса и транспортируется в межмембранное пространство в своей депротонированной форме.Наши первоначальные результаты, которые показывают, что FA может связываться с матриксным сайтом, противоречат результатам экспериментов patch-clip на митопластах, показывая неспособность UCP1 связывать FAs на стороне матрицы [18]. Дальнейшие разработки модели UCP2 проложат путь к прояснению этого вопроса и полного транспортного механизма UCP2.В заключение, мы предлагаем протокол, включающий моделирование гомологии белка UCP2 на основе структуры ANT и последующее микросекундное моделирование молекулярной динамики.Это дает функционально релевантную структуру, которая может быть использована для будущих механистических исследований переноса протонов / ЖК в митохондриях.
4. Материалы и методы
4.1. Детали моделирования
Для анализа структуры белка UCP2, схематически представленной на рисунке S1, мы смоделировали и сравнили две различные структуры белка UCP2 и бычьего белка ANT в качестве эталона. Смоделированные структуры включают (а) опубликованную ЯМР-структуру mUCP2 (код PDB 2LCK, организм Mus musculus) [19], определенную заменой молекулярного фрагмента ЯМР, (b) смоделированную структуру гомологии первичной последовательности белка UCP2 с кристаллографической структурой Белок ANT (код PDB 1OKC, 2.2 Å, высокое разрешение, организм Bos taurus) [38], гомология которого с mUCP2 составляет 24%, и (c) кристаллографическая структура белка ANT, которая служила шаблоном для моделирования гомологии UCP2. Более того, мы использовали его как эталонную структуру для исследуемых структур UCP2. Три различные системы были приготовлены с использованием модуля построения мембран CHARMM-GUI (http://www.charmm-gui.org/) [59,60,61] из трех структур исходного белка UCP2 / ANT, погруженных в 1,2-диолеоил -sn-глицеро-3-фосфохолин (DOPC) мембрана, содержащая 230 молекул липидов, 28 750 молекул воды и 15/19 ионов хлора для нейтрализации общего заряда белка UCP2 / ANT.DOPC был выбран, поскольку он представляет собой один из основных липидов внутренних мембран митохондрий. Смоделированная структура UCP2 и отсутствующие остатки (по сравнению с кристаллографически определенными структурами ANT) — остаток 1 и остатки 294–297 для 1OKC и остатки 1–13 для 2LCK — были добавлены с использованием программы USCF Chimera [62]. Все аргинины и лизины были получены в их протонированных формах, гистидины и цистеины в их нейтральных формах, при этом глутаматы и аспартаты находились в их депротонированных формах.Для всех систем использовалась процедура минимизации и уравновешивания конструктора мембран CHARMM-GUI [59]. После уравновешивания мы смоделировали каждую систему в течение 2,0 мкс с использованием несмещенного моделирования полностью атомной молекулярной динамики (МД) в периодическом прямоугольном прямоугольнике размером 9,5 нм × 9,5 нм × 13,5 нм с шагом по времени 2 фс с силовым полем CHARMM36m [63] и Водная модель TIP3P [64]. Чтобы проверить потенциальный сайт связывания аниона жирной кислоты в UCP2, мы также смоделировали дополнительные четыре системы структур белка UCP2 с добавленным анионом арахидоновой кислоты (AA — ).АК использовался, поскольку он показывает существенное влияние на протонную проводимость через митохондриальные мембраны [10]. AA — был добавлен к сайту связывания аргинина R60 в трех системах: (а) смоделированная структура UCP2 (UCP2 h ), смоделированная через 2 мкс, (b) структура ЯМР после начального уравновешивания, которая очень похожа на экспериментальную ЯМР структура, и (c) экспериментальная структура ЯМР (UCP2 NMR ), смоделированная через 2 мкс. В четвертой системе AA — был добавлен к мутированному сайту связывания (от R60 до S60) в UCP2 h , смоделированном через 2 мкс (UCP2-R60 h ).Каждая из четырех упомянутых выше систем, содержащих AA — , была смоделирована в течение 500 нс и проанализирована дополнительно. Все производственные МД-моделирование проводилось в изобарно-изотермическом ансамбле (NPT) при T = 310 K, который поддерживался с помощью термостата Нозе-Гувера [65,66] независимо для трех групп: DOPC, вода / ионы и белковая подсистема с константа связи 1,0 пс −1 . Давление устанавливали на 1,013 бар и регулировали с помощью полуизотропного баростата Парринелло-Рахмана [67] с постоянной времени для соединения давления 5 пс -1 .Периодические граничные условия (PBC) были наложены во всех трех направлениях, с дальнодействующими электростатическими взаимодействиями, рассчитанными методом Эвальда (PME) [68], с отсечкой кулоновских взаимодействий в реальном пространстве на 1,2 нм с использованием шага Фурье 0,12 нм. и схема отсечки Верле. Все модели были воспроизведены с помощью программного пакета GROMACS 2018.4 [69] и визуализированы с помощью программы VMD (Visualize Molecular Dynamics) [70].4.2. Моделирование гомологии
Для моделированной структуры гомологии мы использовали последовательность из белка 2LCK.Используя моделирование сравнительной структуры белка Blast [71] в UniProt Protein Knowledgebase [72], мы получили наиболее подходящие третичные структуры: 1OKC, 2C3E и 4C9G. Лучшая совпадающая третичная структура 1OKC (наименьшее значение значимости — 4,7 × 10 -20 и наивысшая оценка выравнивания — 88) служит шаблоном структуры. Из выравнивания последовательности мишень-матрица мы создали 10 моделей и выбрали модель с самым низким значением дискретной оптимизированной энергии белка (DOPE) 0,85 в качестве репрезентативной структуры для проведения моделирования.Гомологические модели были построены с помощью программ MODELLER 9 [73] и Chimera 1.13.1 [62].4.3. Расчеты проницаемости
Мы рассчитали осмотическую проницаемость для четырех различных структур UCP2, полученных в результате несмещенного МД моделирования, точно следуя процедуре, проиллюстрированной Zoonens et al. [20] и основан на алгоритме, описанном Аксементьевым и Шультеном [50]. В этом отношении осмотическая проницаемость гомологически смоделированного UCP2 была рассчитана с использованием структур, полученных после 2 мкс несмещенного МД-моделирования, в то время как такое же свойство было определено для трех различных структур ЯМР UCP2, т.е.е., структуры, полученные сразу после уравновешивания, после 0,2 мкс и 2 мкс несмещенного МД моделирования. Четыре выбранные системы были распространены в ансамбле NVT, где позиционные ограничения были применены к атомам C α белка (500 ккал моль -1 нм -2 ). Моделирование распространялось на 10 нс с шагом по времени 2 фс, при этом последние 5 нс использовались в последующем анализе. Моментальные снимки моделирования сохранялись каждые 250 шагов, т.е. каждые 0,5 пс. Условия PBC применялись во всех трех направлениях и обрабатывались электростатикой дальнего действия с использованием метода PME (подробности см. Выше).Пора, образующаяся в структуре как UCP2 NMR , так и UCP2 h , имеет сложную топологию; Таким образом, необходим тщательный выбор области для расчета осмотической проницаемости. Следуя процедуре Zoonens et al. В [20] мы рассчитывали проницаемость с учетом только центральной области поры. Эта область довольно хорошо сохранилась во время моделирования, а ее топология относительно проста, то есть ее можно точно описать как примерно цилиндрическую по своей природе.В этом отношении выбранная область для структур UCP2 ЯМР определялась двумя примерно копланарными кольцами, каждое из которых состоит из шести атомов C α . Каждый атом C α был выбран так, чтобы он принадлежал разной трансмембранной спирали, присутствующей в белке. Выбранные атомы C α образуют нижнее и верхнее кольцо и принадлежат остаткам 34, 85, 137, 181, 239 и 274, а также остаткам 20, 101, 120, 194, 227, 288 соответственно. Из-за разной топологии белка UCP2 h мы использовали аналогичный, но несколько иной выбор атомов C α для определения его центральной поры.В этом отношении нижнее и верхнее кольца были описаны с использованием атомов C α , принадлежащих остаткам 34, 82, 137, 181 и 274, и остаткам 20, 101, 120, 192 и 288, соответственно (см. Рисунок S8). .Таким образом, область, заключенная между двумя выбранными кольцами, имеет форму цилиндра с основаниями в центрах масс нижнего (R 0 ) и верхнего кольца (R 1 ) соответственно. Ось этого цилиндра лежит вдоль вектора R 1 –R 0 . Радиус цилиндра (r = 2 нм) был выбран таким, чтобы он был достаточно большим, чтобы охватить все молекулы воды, обнаруженные в анализируемой поре белка UCP2.
Коллективное смещение молекул воды внутри поры белка в момент времени t + ∆t траектории МД-моделирования теперь можно рассчитать с помощью подхода, разработанного Zhu et al. [74], т. Е. Черезn (t + Δt) = n (t) + ∑i∈S (t, t + Δt) (ΔriΔeL)
где объединение всех подмножеств молекул воды, которые находятся внутри цилиндра в момент времени t и t + ∆t, обозначено как S (t + ∆t), ∆ r i представляет смещение i-й молекулы воды в временное окно от t до t + ∆t, e представляет единичный вектор вдоль R 1 –R 0 .L обозначает длину цилиндрической области и во всех рассмотренных случаях приблизительно равна 1,95 нм. Важно отметить, что смещения молекул воды, которые входят в цилиндрическую область или выходят из нее между двумя последовательными рамками, были вырезаны на границах области таким образом, что учитывалось только смещение таких молекул воды внутри области. Коллективный коэффициент диффузии воды внутри белка, D n , было рассчитано с использованием ⟨n (t)⟩ 2 = 2D n t, при этом среднее значение было получено по 100 субтраекториям, каждая из которых имеет длину 50 пс (5 нс общей время моделирования после уравновешивания / 100), Рисунок S9.Осмотическая проницаемость оценивалась с использованием P f = v w D n , где средний объем одной молекулы воды обозначен как v w . Наконец, полученные таким образом осмотические проницаемости были масштабированы с коэффициентом 1 / 2,87, поскольку реальная вода обладает большей вязкостью по сравнению с используемой моделью воды TIP3P [50].4.4. Связывание АТФ в полости UCP2
Чтобы проверить геометрию сайта связывания АТФ в полости белка UCP2, мы выполнили ряд симуляций (50 на проверенную структуру UCP2).Первоначально мы поместили молекулу АТФ внутрь полости UCP2 для трех различных белковых структур: UCP2 h и двух структур UCP2 ЯМР . Точнее, чтобы представить UCP2 h , мы выбрали структуру, полученную после 2 мкс соответствующей им симуляции свободной МД, в то время как две различные структуры ЯМР UCP2 , а именно структуры, полученные после 20 нс и после 2 мкс их соответствующей свободной МД. моделирование, были использованы для проверки поведения АТФ в полости UCP2 ЯМР .Для каждой структуры UCP2 было выполнено 50 различных моделей MD продолжительностью 20 нс каждое (всего 1 мкс на структуру UCP2). Исходное положение АТФ сохранялось для каждого из 50 моделирования, причем начальные скорости генерировались случайным образом в соответствии с распределением Больцмана. Таким образом, в то время как начальная структура в каждых 50 наборах моделирования (для каждой исследуемой структуры UCP2) представлена одной и той же точкой в конформационном фазовом пространстве, ее положение в импульсном фазовом пространстве отличается, представляя в целом различные исходные структуры.Чтобы проверить, влияют ли начальные условия на полученные результаты, мы провели дополнительное моделирование (снова 20 нс каждое) для структуры UCP2 h , где мы поместили молекулу АТФ в пять разных мест в полости UCP2 h и выполнили 10 моделирования. для каждой стартовой конфигурации. Во всех упомянутых выше симуляциях при анализе использовались только последние 10 нс. Первые 10 нс не учитывались (время уравновешивания). Параметры силового поля CHARMM36m использовали для описания фрагмента АТФ вместе с вышеупомянутыми параметрами липидов UCP2 и DOPC.Моделирование проводилось в ансамбле NVT, при этом все остальные параметры моделирования MD были идентичны тем, которые применялись в моделированиях длительностью 2 мкс (см. Детали моделирования).
4.5. Химические вещества
1,2-диолеоил-sn-глицеро-3-фосфохолин (DOPC), 1,2-диолеоил-sn-глицеро-3-фосфоэтаноламин (DOPE), кардиолипин (CL) из бычьего сердца, арахидоновая кислота (AA), октилполиоксиэтилен Triton X-114, дитиотреитол (DTT), бычий сывороточный альбумин (BSA), аденозин и гуанозинтрифосфат (ATP и GTP), сульфат натрия (Na 2 SO 4 ), (гидрофосфат диаммония NH 4 ) 2 HPO 4 ), 2- (N-морфолино) этансульфоновая кислота (MES), 2-амино-2- (гидроксиметил) пропан-1,3-диол (Трис), этиленгликоль- бис (β-аминоэтиловый эфир) -N, N, N ‘, N’-тетрауксусная кислота (EGTA), гексан, гексадекан и додецилсульфат натрия (SDS) были получены от Sigma-Aldrich (Мюнхен, Германия).Хлороформ был от Merck KGaA (Дармштадт, Германия).
4.6. Клонирование, мутация и экспрессия mUCP2 и реконструкция в липосомы.
UCP2 мыши (mUCP2) был клонирован и экспрессирован, как описано ранее [75,76]. Вкратце, ORF mUCP2 была вставлена в плазмиду экспрессии pET24a-. Для экспрессии белка плазмиду mUCP2 дикого типа переносили в клетки E. coli (штамм Rosetta) и выращивали до достижения OD 600 нм между 0,3 и 0,5. Затем экспрессию белка индуцировали добавлением 1 мМ изопропил-β-D-тиогалактопиранозида (IPTG).Клетки E. coli инкубировали в течение 3 ч перед сбором центрифугированием. Тельца включения (IB), содержащие экспрессированные белки, собирали путем разрушения клеток с помощью французского пресса после центрифугирования при 14000 × g [77].Сайт-направленный мутагенез in vitro проводили на экспрессионных плазмидах, содержащих кДНК mUCP2 в качестве матриц. Мутация была введена с помощью сконструированного олигонуклеотида для изменения R60 (CGT) на S (AGT) с использованием набора для сайт-направленного мутагенеза Q5 (New England Biolabs GmbH, Франкфурт-на-Майне, Германия) и подтверждена секвенированием.
Рекомбинантные mUCP2WT и mUCP2R60S были очищены и повторно свернуты из телец включения и преобразованы в липосомы в соответствии с ранее описанным протоколом [54]. Вкратце, mUCP3 солюбилизировали из IB с использованием 2% саркозила и 1 мМ DTT. К солюбилизированному mUCP2 добавляли 100 мг полярного липида E.coli (полярные липиды Avanti, Алабастер, штат Алабама, США), 300 мкг Triton X-114, 75 мкг октил-полиоксиэтилена и 2 мМ GTP. Саркозил и GTP удаляли диализом с буфером для анализа (50 мМ Na 2 SO 4 , 10 мМ Трис, 10 мМ MES и 0.6 мМ EGTA, pH 7,34). Образец пропускали через колонку с гидроксиапатитом (Bio-Rad, Laboratories, Inc., Фельдкирхен, Германия) для удаления разложившихся белков. Неионные детергенты удаляли с помощью Bio Beads (Bio-Rad, Германия). Чистота рекомбинантных белков была подтверждена окрашиванием серебром (рис. S10). Правильное сворачивание было доказано анализом активности — активация или ингибирование белка.4.7. Измерения электрических параметров мембран, восстановленных с помощью mUCP2
Плоские липидные бислои были сформированы из (протео-) липосом [78,79], состоящих из 45:45:10 мол.% DOPC: DOPE: CL. Концентрация липидов составляла 1,5 мг / мл, а соотношение белков и липидов — 4 мкг на мг липидов. Арахидоновую кислоту (АК) в концентрации 15 мол.% Добавляли непосредственно в липидную фазу перед формированием мембраны. Буфер содержал 50 мМ Na 2 SO 4 , 10 мМ Трис, 10 мМ MES и 0,6 мМ EGTA при pH = 7,34 и T = 306 К. Правильное образование мембраны было проверено путем измерения удельной емкости (C = 718 ± 34 нФ. / см 2 ), который не зависел от содержания белка, АК и АТФ.Измерения ток-напряжение (I-U) выполняли с помощью усилителя с коммутационным зажимом (EPC 10 USB, HEKA Elektronik Dr Schulze GmbH, Ламбрехт, Германия). Общая проводимость мембраны при 0 мВ была получена из наклона линейной аппроксимации экспериментальных данных при приложенных напряжениях от -50 мВ до +50 мВ (рисунок S7). АТФ растворяли в буферном растворе до концентрации 400 мМ и pH исходного раствора доводили до 7,34. Объем 3,75 мкл исходного раствора добавляли к 750 мкл буферного раствора до конечной концентрации 2 мМ АТФ.Время инкубации составляло 30 мин при Т = 306 К. Данные анализировали с использованием Sigma Plot (Systat Software GmbH, Эркрат, Германия).Развязка глобальных детерминант времени репликации и точная настройка перицентрического гетерохроматина мыши | Журнал клеточной биологии
Хромоцентры мыши представляют собой кластеры поздно реплицирующегося перицентрического гетерохроматина, содержащего HP1, связанный с триметилированным лизином 9 гистона h4 (Me 3 K9h4). Используя бесклеточную систему для инициации репликации в ядрах G1-фазы, мы демонстрируем, что хромоцентры приобретают свойство поздней репликации, совпадающей с их реорганизацией после митоза и установлением глобальной программы синхронизации репликации.HP1 диссоциирует во время митоза, но восстанавливается до установления поздней репликации, и удаление HP1 из хромоцентров за счет конкуренции с пептидами Me 3 K9h4 не приводит к ранней репликации, демонстрируя, что это взаимодействие не является ни необходимым, ни достаточным для поздней репликации. Однако в клетках, лишенных метилтрансфераз Suv39h2,2, ответственных за триметилирование K9h4 и связывание HP1 в хромоцентрах, репликация хромоцентровой ДНК была продвинута на 10-15% длины S фазы.Повторное введение активности Suv39h2 восстановило более позднее время репликации. Мы пришли к выводу, что активность Suv39 необходима для тонкой настройки репликации перицентрического гетерохроматина относительно др. Поздно реплицирующихся доменов, тогда как отдельные факторы устанавливают глобальную временную программу репликации во время ранней фазы G1.
В клетках млекопитающих репликация ДНК происходит в определенном пространственно-временном порядке. В общем, эухроматические домены располагаются внутри ядра и реплицируются в ранней S фазе, тогда как гетерохроматические домены локализуются на периферии ядра или около ядрышек и реплицируются поздно (Schwaiger and Schubeler, 2006; Zink, 2006).Хотя эта пространственно-временная организация была оценена в течение некоторого времени, ее функциональное значение не понято. Поскольку хроматин собирается на вилке репликации, временная сегрегация может предоставить важную регуляторную возможность (McNairn and Gilbert, 2003). Действительно, репортерные гены, микроинъектированные в ядра млекопитающих в разное время, собираются в разные типы хроматина (Zhang et al., 2002), а время репликации и субядерное положение некоторых генов регулируется в процессе развития (Hiratani et al., 2004; Перри и др., 2004; Williams et al., 2006).
Путем введения ядер G1-фазы в бесклеточную систему репликации мы ранее продемонстрировали, что программа времени репликации устанавливается в дискретной точке во время ранней фазы G1, называемой точкой выбора времени (TDP; Dimitrova and Gilbert, 1999; Li et al. ., 2001, 2003). Интересно, что внутриядерное пространственное репозиционирование хромосомных доменов, а также кластеризация синхронно запускающихся источников репликации происходит в течение того же самого короткого промежутка времени.Сходное событие в ранней G1-фазе может регулировать положение субядра и время репликации у почкующихся дрожжей (Raghuraman et al., 1997; Heun et al., 2001a). Что не было ясно, так это то, восстанавливается ли программа репликации конститутивного гетерохроматина в каждом клеточном цикле. В этом исследовании мы изучили установление поздней репликации перицентрического гетерохроматина мыши.
Хромоцентры содержат большое центральное ядро перицентрического гетерохроматина, состоящее из повторов γ-сателлитной ДНК, упакованных в хроматин, который содержит гистон h4, триметилированный по лизину 9 (Me 3 K9h4; Peters et al., 2003; Райс и др., 2003). Триметилирование, которое осуществляется гистоновыми метилтрансферазами Suv39h2,2, создает сайт связывания с высоким сродством (хотя и зависит от контекста) для HP1α и -β, которые концентрируются внутри хромоцентров (Bannister et al., 2001; Lachner et al., 2001; Стюарт и др., 2005). Локализация HP1 в хромоцентрах является логическим кандидатом на роль детерминанты времени репликации, учитывая параллели между белками HP1 и белками молчащего хроматина (Sir) у почкующихся дрожжей (Jones et al., 2000; Wang et al., 2000). Белки Sir концентрируются в кластерах теломер, прикрепленных к периферии ядра. Кластеризация теломер создает сток для белков Sir, изолируя их на периферии и предотвращая их замалчивание активного генома (Gartenberg et al., 2004). Теломеры реплицируются поздно, и белки Sir необходимы для поздней репликации некоторых источников репликации дрожжей (Stevenson and Gottschling, 1999; Zappulla et al., 2002).
Мы обнаружили, что программа времени репликации хромоцентров восстанавливается одновременно с реорганизацией перицентрического гетерохроматина в хромоцентры.Таким образом, TDP влияет одновременно на многие типы хроматина. Однако Suv39h2,2-опосредованное триметилирование K9h4 и взаимодействие HP1 с хроматином не были ни необходимыми, ни достаточными для установления поздней репликации на TDP. Вместо этого Suv39 необходим для частичной задержки репликации хромоцентров относительно др. Поздно реплицирующихся доменов, демонстрируя, что глобальная программа синхронизации устанавливается независимо от дополнительных факторов, которые точно настраивают программу репликации.
Хромоцентры мыши реплицируются во второй половине S фазы (Guenatri et al., 2004; Wu et al., 2005). Чтобы определить, устанавливается ли время репликации хромоцентров во время ранней фазы G1, мышиные клетки C127 синхронизировались в митозе и высвобождались в фазу G1 в течение разных периодов времени (G1 1, 2 и 3 часа). Интактные ядра вводили в экстракты яиц Xenopus laevis , и ДНК, синтезированную in vitro, подвергали импульсной метке биотин-dUTP в разное время после инициации репликации.FISH с мышиным γ-сателлитным (основным сателлитным) ДНК-зондом использовали для визуализации перицентрического гетерохроматина, а совместную локализацию сигналов FISH с реплицированной ДНК, которая была идентифицирована путем окрашивания меченым авидином, отслеживали как индикатор репликации хромоцентра (рис. 1). В 1- и 2-h ядрах G1 хромоцентры реплицировались при самых ранних обнаруживаемых признаках синтеза ДНК, что указывает на отсутствие временной специфичности. Напротив, репликация хромоцентров в 3-h ядрах G1 была значительно задержана, указывая на то, что программа поздней репликации для хромоцентров была установлена между 2 и 3 часами после митоза.Поскольку общая скорость репликации была идентична для всех трех популяций ядер (рис. 1 d), ранняя репликация хромоцентров в 1- и 2-h ядрах G1 не была просто вызвана повышенной скоростью синтеза ДНК.
Это временное окно (2–3 часа после митоза) позже, чем ранее охарактеризованный TDP в клетках CHO (∼1–2 часа после митоза; Dimitrova and Gilbert, 1999; Li et al., 2001, 2003). Чтобы определить, было ли это различие результатом специфичных для типов клеток различий в глобальном TDP или явно более позднего установления времени репликации конститутивного гетерохроматина, мы оценили глобальную пространственно-временную программу репликации в C127 (рис.2). Асинхронно растущие клетки были помечены импульсом BrdU и отслеживались в течение периода времени (4–5 ч), который был оптимизирован для получения наивысшего процента митотических клеток в поздней S-фазе во время импульса BrdU (около 50% митотических фигур отображают метку BrdU. , с> 95% метки из поздней фазы S). Ядра из клеток, высвобожденных в фазу G1 в течение 1, 2 или 3 ч, вводили в экстракт яиц Xenopus , а участки самого раннего синтеза ДНК in vitro (первые 30 мин) отслеживали с помощью включения биотин-дУТФ (рис.2 б). Совместная локализация поздно реплицирующейся метки BrdU с меченным биотином ранним синтезом ДНК in vitro указывает на отсутствие временной специфичности. В этих экспериментах присутствие или отсутствие колокализации можно было четко определить вручную с помощью двойного (красный / зеленый) фильтра (оценка для желтой окраски), что устраняет необходимость в громоздком анализе колокализации, представленном на рис. 1. Это различие может быть вызвано за счет улучшенного сохранения трехмерной структуры в ядрах, которые не были денатурированы для анализа FISH.Как показано на рис. 2c, большинство G1 1- и 2-часовых, но очень мало 3-часовых ядер демонстрируют желтые фокусы, демонстрируя, что общая программа времени репликации в клетках C127 мыши устанавливается между 2 и 3 часами после митоза. Это более позднее время фазы G1 для TDP в клетках C127 по сравнению с клетками CHO может быть результатом более длительной фазы G1 на 2 часа в этих клетках по сравнению с клетками CHO (неопубликованные данные).
Чтобы определить время, в которое хромосомные домены репозиционируются после митоза, поздняя метка S BrdU отслеживалась в ядрах G1.Поскольку большой процент синтеза ДНК в поздней S-фазе происходит на периферии ядра, мы количественно оценили процент ядер, которые репозиционировали метку BrdU на периферию ядра. Как показано на рис. 2d, с ядрами G1 1- и 2-h <10% BrdU-положительных клеток отображали метку в виде периферического рисунка, тогда как этот процент достигал почти 50% (уровень плато, эквивалентный уровню, полученному при гораздо более поздние периоды клеточного цикла) через 3 часа после митоза. Мы пришли к выводу, что хотя разные типы клеток могут восстанавливать время репликации в несколько разное время после митоза, TDP, тем не менее, совпадает с репозиционированием доменов хромосом и одновременно влияет на многие разные типы хроматина.
Хотя большие блоки перицентрической AT-богатой сателлитной ДНК легко видны при окрашивании FISH или DAPI на протяжении всего клеточного цикла, мы тем не менее можем наблюдать их реорганизацию в структуры более правильной формы во время перехода TDP (Fig. 3a). Поскольку триметилирование K9h4 и связывание белков HP1 участвуют в сборке гетерохроматина (Grewal and Rice, 2004), мы исследовали присутствие белков Me 3 K9h4 и HP1 в хромоцентрах, когда клетки проходят через TDP.Me 3 K9h4 был сконцентрирован в DAPI-плотных областях до TDP (рис. 3 a), и общее количество Me 3 K9h4 в клетках не изменилось в течение этого времени (рис. 3 b). Поскольку мы и другие показали, что Me 3 K20h5 также обогащен перицентрическим гетерохроматином (Kourmouli et al., 2004; Schotta et al., 2004), и некоторые исследования предполагают, что эта модификация может регулироваться клеточным циклом (Fang et al. , 2002; Rice et al., 2002), мы также исследовали его численность во время TDP, но изменений не обнаружено.Следовательно, этих двух модификаций гистонов в перицентрическом гетерохроматине недостаточно для установления поздней репликации.
Предыдущие эксперименты на Drosophila melanogaster (Pak et al., 1997) и клетках млекопитающих (Fischle et al., 2005; Hirota et al., 2005) продемонстрировали, что большая часть HP1 диссоциирует от хроматина во время митоза и реассоциируется впоследствии, делает HP1 интересным кандидатом на роль белка, участвующего в сбросе времени репликации на TDP.Мы подтвердили как иммунофлуоресценцию, так и визуализацию живых клеток GFP-меченных белков HP1, что HP1α и -β были в значительной степени рассредоточены во время митоза (неопубликованные данные). Однако все детектируемые HP1α и -β отскакивают к хроматину за счет анафазы (не изображены) и можно ясно видеть сконцентрированными в DAPI-плотных областях в ядрах G1 1-h (рис. 3 a). Чтобы определить, совпадает ли какое-либо изменение аффинности белков HP1 к хроматину с TDP, мы экстрагировали растворимые клеточные белки из клеток до и после TDP с неионным детергентом при различных концентрациях соли (рис.3, в, г). Эти результаты показали, что примерно половина белков HP1 растворима или легко диссоциирует от метафазного хроматина. Однако через 1 час после митоза все определяемые HP1α и -β были очень тесно связаны с хроматином без обнаруживаемого изменения аффинности в TDP (рис. 3 d). Мы пришли к выводу, что взаимодействие HP1-Me 3 K9h4 в перицентрическом гетерохроматине происходит до TDP и, следовательно, недостаточно для установления программы времени поздней репликации хромоцентров.
Чтобы выяснить, необходима ли ассоциация HP1 для поздней репликации, мы воспользовались бесклеточной природой нашей системы, чтобы удалить HP1 из хроматина после TDP перед репликацией in vitro с использованием пептида, имитирующего метилированный хвост h4 (Bannister et al., 2001). Ядра из клеток, синхронизированных через 3 ч после митоза, инкубировали с триметилированным пептидом, состоящим из первых 20 аминокислот гистона h4 (рис.4 а). В качестве контроля аликвоты одних и тех же ядер инкубировали либо с неметилированной формой того же пептида, либо без пептида. Инкубация с триметилированным, но не с контрольным пептидом привела к солюбилизации 30-40% общего белка HP1 (рис. 4b) и удалению почти всего детектируемого HP1 в хромоцентрах (рис. 4c). Фактически, HP1 оставался связанным с хроматином, окружающим хромоцентры, но селективно удалялся из самих DAPI-плотных хромоцентров. Это указывает на то, что Me 3 K9h4 является первичным сайтом связывания HP1 в хромоцентрах, тогда как HP1 в других сайтах связывается с другими компонентами хроматина, которые, как известно, связывают HP1 (Polioudaki et al., 2001; Сингх и Георгатос, 2002). Эти ядра затем вводили в экстракт яиц Xenopus и оценивали совместную локализацию самого раннего синтеза ДНК in vitro с γ-сателлитной ДНК. Истощение HP1 в хромоцентрах не оказывало значительного влияния на синхронизацию этих доменов или общую скорость синтеза ДНК in vitro (рис. 4, d и e).
Вышеупомянутые эксперименты демонстрируют, что взаимодействия HP1-Me 3 K9h4 недостаточно для установления поздней репликации хромоцентров.Это было неожиданно в свете недавних связей между структурой хроматина и временем репликации как в системах млекопитающих (Li et al., 2004; Takebayashi et al., 2005), так и в дрожжевых системах (см. Обзор Donaldson, 2005), а также ролью белков HP1 в образование перицентрического гетерохроматина (Maison, Almouzni, 2004). Чтобы выяснить, играет ли это взаимодействие какую-либо роль во времени репликации хромоцентров, мы исследовали репликацию хромоцентров в клетках, лишенных Suv39h2 и Suv39h3 (Peters et al., 2001). Эмбриональные фибробласты мыши (MEF), полученные от мышей, лишенных обоих этих ферментов (двойной нуль; Suv39dn), имеют <30% от общего количества клеточного Me 3 K9h4 и не обнаруживают Me 3 K9h4 в хромоцентрах (Peters et al., 2003; Райс и др., 2003). В этих клетках белки HP1 остаются прочно связанными с хроматином, но истощаются из хромоцентров (рис. S1). Это согласуется с результатами на рис. 4, показывающими, что HP1 привязан к хромоцентрам через Me 3 K9h4, но связывается с др. Сайтами хроматина с помощью других механизмов. Повторное введение активности Suv39 путем стабильной трансфекции экспрессирующим вектором Suv39h2 (Rice et al., 2003) частично восстанавливает Me 3 K9h4 и HP1 в хромоцентрах (рис. S2), обеспечивая «дополнительный» контроль для проверки наличия любых различий являются результатом потери активности Suv39.
Для определения времени репликации хромоцентров в этих трех клеточных линиях мы использовали метод ретроактивной синхронизации, который обычно используется для анализа времени репликации специфических генных последовательностей (Hiratani et al., 2004). Этот метод устраняет необходимость в громоздких методах синхронизации, специфичных для клеточной линии, которые могут нарушить клеточный цикл. После импульсного мечения растущей ДНК с помощью BrdU клетки ретроактивно сортировали с помощью проточной цитометрии на популяции на разных стадиях S-фазы (рис.5 а). Геномную ДНК выделяли из каждой фракции, и возникающую (замещенную BrdU) ДНК иммунопреципитировали антителами против BrdU. Аликвоты этих препаратов формирующихся цепей были иммобилизованы на нейлоновых фильтрах и гибридизированы с зондами, содержащими либо основные, либо второстепенные повторы сателлитной ДНК (Рис. 5b), которые характеризуют перицентрическую и центромерную ДНК, соответственно (Peters et al., 2003). В качестве контроля мы наблюдали за репликацией генов α- и β-глобина (не показаны), которые реплицируются рано и поздно, соответственно, и митохондриальной ДНК (рис.5 b), который реплицируется на протяжении всего клеточного цикла и в равной степени представлен в образующихся препаратах ДНК из всех времен клеточного цикла (Bogenhagen and Clayton, 1977; James and Bohman, 1981; Magnusson et al., 2003). Как показано на рис. 5c, второстепенная сателлитная ДНК реплицировалась во время фазы S значительно раньше, чем основная сателлитная, но мы не смогли обнаружить значительных различий в программе репликации этих последовательностей ДНК ни в Suv39dn MEF, ни в спасенных дополнительных клетках. линия.
Молекулярные анализы на рис.5 подтверждают исследования in vitro на рис. 4 и демонстрируют, что взаимодействие Suv39h2,2 и HP1-Me 3 K9h4 не является ни необходимым, ни достаточным для поздней репликации перицентрического гетерохроматина. Однако мы действительно заметили небольшое, но не статистически значимое увеличение времени репликации основной сателлитной ДНК в D15 (рис. 5 c). Небольшие изменения во времени репликации лучше выявляются клеточными анализами. Например, небольшие различия во времени репликации импринтированных генов и генов иммуноглобулинов могут быть обнаружены с помощью клеточных анализов (Mostoslavsky et al., 2001; Singh et al., 2003), но не молекулярный анализ (Zhou et al., 2002). Таким образом, мы оценили время репликации хромоцентров в индивидуальных клетках, используя метод отслеживания импульсов, который также не требует синхронизации клеток (Wu et al., 2005). Клетки были помечены импульсным способом 5′-хлор-2′-дезоксиуридином (CldU), отслеживались в течение различных промежутков времени, а затем импульсной меткой 5′-йод-2′-дезоксиуридином (IdU). Сайты включения CldU и IdU были обнаружены с помощью иммунофлуоресценции с использованием CldU- и IdU-специфических антител (Aten et al., 1992). Изменение в распределении этих двух меток в отдельных ядрах после разного времени погони показывает временной порядок этих паттернов репликации. Примерные объединенные изображения CldU и IdU в MEF Suv39dn показаны на фиг. 6a. Общее пространственное расположение и временной порядок типичных шести паттернов репликации мышей (Wu et al., 2005), которые подробно описаны на рис. 6b, в значительной степени не изменились между MEF дикого типа и Suv39dn, что демонстрирует, что Me 3 K9h4 не требуется для поддержания общей программы пространственно-временной репликации.Однако тщательное изучение паттернов в MEF Suv39dn показало, что истинный паттерн III не наблюдался (рис. 6b). Скорее, казалось, что репликация хромоцентров (обычно специфическая для паттерна IV) уже имела место в клетках, которые в остальном были характерны для паттерна III.
Чтобы конкретно оценить время репликации хромоцентра относительно глобальной программы репликации, мы рассчитали время, необходимое клеткам для перехода от пространственных паттернов репликации, характерных для ранней S-фазы, до времени репликации хромоцентров (рис.6, в, г). Это было количественно определено как процент ядер, которые имели CldU-меченные ранние S-фазные паттерны и IdU-меченные хромоцентры в разное время преследования. Эти результаты показали, что репликация хромоцентров была продвинута в MEF Suv39dn по сравнению с MEFs дикого типа на 10-15% S фазы. Повторное введение активности Suv39 в клеточную линию add-back в основном восстанавливает эту небольшую задержку репликации хромоцентра (рис. 6, c и d).
В ходе экспериментов, описанных на рис.6, мы распознали присутствие заметного тела хроматина, который был интенсивно помечен BrdU во время синтеза ДНК паттерна III и реплицировался синхронно с хромоцентрами только в мутантных MEFs Suv39dn (рис. 6). Это тело также интенсивно окрашивалось антителом, специфичным к триметилированному лизину 27 гистона h4 (рис. 7а), модификации, которая сильно обогащена инактивированной поздно реплицирующейся Х-хромосомой (Plath et al., 2003; Silva et al., 2003 г.). Учитывая, что эти MEF произошли от эмбриона самки мыши, весьма вероятно, что это тело является поздно реплицирующейся неактивной Х-хромосомой (Xi), и для целей обсуждения мы будем называть ее Xi.Важно, что в MEF дикого типа (рис. 7, b и d) и в MEF Suv39dn, спасаемых с помощью добавления Suv39h2 (рис. 7 d), репликация хромоцентра имеет место отчетливо после репликации Xi. Напротив, Suv39dn MEFs реплицируют хромоцентры одновременно с репликацией Xi (рис. 7, c и d) в 75% Xi-меченных клеток. Поскольку Xi не обогащен Me 3 K9h4 (Kohlmaier et al., 2004), можно видеть, что хромоцентры реплицируются в другом паттерне III вместе с последними остатками небольших внутренних эухроматических фокусов (рис.6 б), мы интерпретируем эти результаты как прогресс репликации хромоцентров, а не задержку репликации Xi. Эти результаты обеспечивают внутренне контролируемый эталон и вместе с данными на рис. 6 демонстрируют, что Suv39-обеспечиваемое триметилирование h4K9 отвечает за задержку времени репликации на 10-15%.
Наши результаты демонстрируют, что задержка репликации перицентрического гетерохроматина налагается в дискретной точке во время ранней фазы G1, что совпадает с субядерной организацией хромоцентров и согласуется с существованием глобального TDP (Dimitrova and Gilbert, 1999; Li и другие., 2001, 2003). Это первая демонстрация TDP для конститутивного гетерохроматина и предполагает, что общее время репликации для всех доменов хроматина устанавливается одновременно. Тесная временная связь между установлением поздней репликации и повторной сборкой субядерных доменов в различных клеточных линиях и у почкующихся дрожжей (Raghuraman et al., 1997; Heun et al., 2001a, b) указывает на некоторый аспект ядерной реорганизации как репликации определитель времени. В этом исследовании мы показываем, что эта детерминанта устойчива к обширным изменениям в структуре хроматина.
HP1 является основным хроматиновым компонентом хроматина хромоцентра. Поскольку мы не смогли обнаружить никаких изменений сродства HP1 к хроматину в это время, детерминанта TDP должна быть независимой от взаимодействия HP1-Me 3 K9h4. Более того, потеря HP1 из хромоцентров у мутантов Suv39dn не приводит к сдвигу репликации хромоцентров к ранней репликации S-фазы, но вызывает частичное улучшение времени репликации.Метилирование ДНК, преобладающая модификация основной сателлитной ДНК, также вряд ли может объяснить раннее установление времени репликации в G1-фазе, поскольку эта ковалентная модификация ДНК не удаляется во время митоза, а мутанты Suv39dn значительно снижают метилирование ДНК в основной сателлитной ДНК (Lehnertz и др., 2003). Наконец, мутанты Suv39dn не показали каких-либо явных изменений в количестве или общей структуре собранных хромоцентров (неопубликованные данные). Вместе эти результаты подтверждают, что некоторые аспекты структуры субядерных доменов хромоцентров необходимы для их общей поздней репликации и собираются в TDP, но это событие не зависит от метилирования ДНК, триметилирования K9h4 и h5K20 и белков HP1.Вместо этого эти основные компоненты хроматина отвечают за тонкую настройку времени репликации по сравнению с другими доменами, обнаруживая по крайней мере два уровня контроля над временем репликации.
Наши результаты демонстрируют, что центромеры мышей реплицируются раньше, чем фланкирующий гетерохроматин. В соответствии с этими результатами, более ранняя репликация центромерного хроматина относительно фланкирующего гетерохроматина наблюдалась в клетках Drosophila (Ahmad and Henikoff, 2001; Sullivan and Karpen, 2001), и было показано, что центромеры человека реплицируются асинхронно с фланкирующим гетерохроматином (Blower и другие., 2002; Салливан и Карпен, 2004). Однако наши результаты противоречат недавнему исследованию, в котором сообщается, что центромерный хроматин мыши реплицируется после перицентрического хроматина (Guenatri et al., 2004), которое было основано на совместной локализации BrdU с основными и второстепенными сателлитными ДНК-зондами. В наших руках интерпретировать результаты с помощью этого метода было очень сложно. Поскольку хромоцентры содержат более одной центромеры, а разные центромеры реплицируются в разное время в течение 2 часов (Wu et al., 2005), иммунофлуоресцентные подходы могут выявить случаи, в которых перицентрический гетерохроматин одной хромосомы реплицируется перед центромерным хроматином другой хромосомы. хромосома в одном хромоцентре.Фактически, более недавнее исследование обнаружило, что хроматин, содержащий кинетохорные белки мыши, реплицируется раньше и шире на протяжении S фазы, чем перицентрический гетерохроматин (Weidtkamp-Peters et al., 2006), что согласуется с нашим молекулярным анализом (Fig. 5).
АктивностьSuv39 непосредственно отвечает за триметилирование K9h4 и ассоциацию HP1 с хромоцентрами, а также требуется для метилирования ДНК (Lehnertz et al., 2003) и триметилирование h5K20 (Kourmouli et al., 2004; Schotta et al., 2004) в перицентрическом гетерохроматине. Таким образом, удивительно, что мутация с таким сильным влиянием на структуру хроматина должна иметь только дополнительный эффект на время репликации. Однако это возрастающее различие похоже на различия во времени между гомологами импринтированных и иммуноглобулиновых генов (Mostoslavsky et al., 2001; Singh et al., 2003), и интригует предположение, что этот дифференциал репликации может вносить вклад в идентичность центромер.Центромеры и перицентрический гетерохроматин имеют совершенно разные белковые составы (Sullivan and Karpen, 2004). Более того, ни HP1, ни Me 3 K9h4 не присутствуют в центромерном хроматине (Lehnertz et al., 2003; Peters et al., 2003), указывая тем самым, что Suv39 не модифицирует центромерный хроматин. Наши результаты предполагают, что разница во времени репликации перицентрического хроматина относительно центромерного хроматина может быть уменьшена у мутантов Suv39h2,2. Более того, мыши Suv39dn демонстрируют нарушенную жизнеспособность, а те, что выживают, имеют задержку роста и повышенный риск опухоли (Peters et al., 2001). Клетки этих мышей обнаруживают серьезные хромосомные аномалии, указывающие на нарушения функции центромеры (Peters et al., 2001; Guenatri et al., 2004). Таким образом, наши результаты повышают вероятность того, что относительное время репликации центромеры и фланкирующего хроматина может быть важным для структуры и функции центромеры.
Мышиные клетки C127 культивировали и синхронизировали в митозе, как описано ранее (Gilbert and Cohen, 1987; для обзора см. Wu et al., 1997). MEF культивировали, как описано ранее (Peters et al., 2001), за исключением 10% FBS. В этих условиях продолжительность S-фазы (измеренная, как показано на рис. 6) составляла 7,2–9,4 ч для дикого типа, 8,6–10,2 ч для D15 и 9–9,7 ч для D15 + Suv39h2. Клеточная линия D15, спасенная Suv39h2, была предоставлена Дж. Райсом (Университет Южной Калифорнии, Лос-Анджелес, Калифорния).
Время репликации хромоцентра. Интактные ядра получали из клеток C127 в G1-фазе и вводили в экстракт яиц Xenopus , как описано ранее (Wu et al., 1997; Dimitrova and Gilbert, 1998). Где указано, ядра инкубировали с пептидами в описанном транспортном буфере, содержащем 1,5% BSA. Пептиды (Cowell et al., 2002) растворяли в дистиллированной воде при концентрации 10 мг / мл, и аликвоты исходных материалов хранили при -70 ° C. В каждый момент времени отбирали аликвоты реакции и импульсно метили 50 мкМ биотин-16-dUTP (Sigma-Aldrich) в течение 5-10 мин.Реакции останавливали разбавлением 1:10 холодным буфером для изоляции ядер, ядра прикрепляли к предметным стеклам и фиксировали, как описано ранее (Li et al., 2001). Перицентрический гетерохроматин был обнаружен с помощью FISH с помощью зонда γ-сателлитной ДНК, а включение биотина-дУТФ было обнаружено с помощью конъюгированного стрептавидина Texas red (GE Healthcare). Γ-сателлитная плазмида, содержащая восемь копий сателлитного повтора длиной 234 п.н. (Lundgren et al., 2000), была помечена ник-трансляцией дигоксигенин-11-dUTP (Life Technologies), и FISH был выполнен, как описано ранее (Li et al. ., 2001).
Асинхронно растущие клетки метили 15 мкг / мл BrdU в течение 30 мин и инкубировали в среде, содержащей 50 нг / мл нокодазола, в течение 4–5 ч перед митотическим встряхиванием. В указанные моменты времени фазы G1 интактные ядра инкубировали в экстракте яиц Xenopus . В различные моменты времени in vitro аликвоты ядер удаляли и пульсируемо метили биотин-dUTP, а совместную локализацию BrdU и биотина определяли, как описано ранее (Dimitrova and Gilbert, 1999).
Процент реплицированной входной ДНК определяли в экстрактах яиц Xenopus с добавлением α- [ 32 P] dATP путем кислотного осаждения, как описано ранее (обзор см. Wu et al., 1997). Обратите внимание, что скорость синтеза ДНК in vitro выше, чем in vivo, из-за более синхронного срабатывания источников репликации, что приводит к сжатой S-фазе, которая, тем не менее, следует тому же временному порядку (Dimitrova and Gilbert, 1998; Li et al., 2003).
МечениеBrdU, сортировку клеток и иммунопреципитацию BrdU-меченой ДНК проводили, как описано ранее (Hiratani et al., 2004). Меченная BrdU ДНК из равного числа клеток была иммобилизована на нейлоновых мембранах и гибридизирована с основной (предоставленной N. Dillon, Совет медицинских исследований, Центр клинических наук, Лондон, Великобритания; Lundgren et al., 2000) и второстепенной (pCR4 Min5). -1; предоставлено Т.Jenuwein, Исследовательский институт молекулярной патологии, Вена, Австрия; Lehnertz et al., 2003) сателлитные ДНК-зонды, а также митохондриальный зонд мыши (p501-1; предоставлен T. Brown, National Institutes of Health, Bethesda, MD; Brown and Clayton, 2002). Зонды маркируют методом случайного прайминга (Invitrogen). Мембраны были гибридизованы и промыты, как описано ранее (для обзора см. Wu et al., 1997), и относительные числа в минуту были получены с помощью анализа фосфорного изображения (Molecular Dynamics).Значения для основной и второстепенной сателлитной ДНК были нормализованы к значениям для гибридизации митохондриальной ДНК, и относительный сигнал гибридизации был представлен как процент от суммы этих нормализованных значений по всем фракциям клеточного цикла.
Мы посвящаем эту рукопись памяти Джона В. Ньюпорта, скончавшегося во время ее подготовки. Мы благодарим T. Jenuwein за антитела Meh4 и pCR4, N.Dillon для основной плазмиды сателлитной ДНК, T. Brown для p501-1 и J. Rice для линии клеток D15, спасенных Suv39h2. Мы также благодарим Ф. Ли, А. Макнэрна, И. Хиратани, Т. Йокочи, Дж. Лу, Б. Нокса и П. Кейна за полезные обсуждения, а также Дж. Чена, Н. Гончорова и Национальный научный фонд (NSF) Студентам REU А. Терри, Дж. Аффортунато, Дж. Транчеллу, М. Пэннингу и Б. Ниссену за техническую поддержку.
Эта работа была поддержана грантом NSF MCB-0077507 и грантом GM-57233 Национального института здравоохранения Д.М. Гилберт.
Этот сайт использует файлы cookie для повышения производительности. Если ваш браузер не принимает файлы cookie, вы не можете просматривать этот сайт.
Настройка вашего браузера для приема файлов cookie
Существует множество причин, по которым cookie не может быть установлен правильно. Ниже приведены наиболее частые причины:
- В вашем браузере отключены файлы cookie. Вам необходимо сбросить настройки своего браузера, чтобы он принимал файлы cookie, или чтобы спросить вас, хотите ли вы принимать файлы cookie.
- Ваш браузер спрашивает вас, хотите ли вы принимать файлы cookie, и вы отказались. Чтобы принять файлы cookie с этого сайта, используйте кнопку «Назад» и примите файлы cookie.
- Ваш браузер не поддерживает файлы cookie. Если вы подозреваете это, попробуйте другой браузер.
- Дата на вашем компьютере в прошлом. Если часы вашего компьютера показывают дату до 1 января 1970 г., браузер автоматически забудет файл cookie. Чтобы исправить это, установите правильное время и дату на своем компьютере.
- Вы установили приложение, которое отслеживает или блокирует установку файлов cookie. Вы должны отключить приложение при входе в систему или уточнить у системного администратора.
Почему этому сайту требуются файлы cookie?
Этот сайт использует файлы cookie для повышения производительности, запоминая, что вы вошли в систему, когда переходите со страницы на страницу. Чтобы предоставить доступ без файлов cookie потребует от сайта создания нового сеанса для каждой посещаемой страницы, что замедляет работу системы до неприемлемого уровня.
Что сохраняется в файлах cookie?
Этот сайт не хранит ничего, кроме автоматически сгенерированного идентификатора сеанса в cookie; никакая другая информация не фиксируется.
Как правило, в файле cookie может храниться только информация, которую вы предоставляете, или выбор, который вы делаете при посещении веб-сайта. Например, сайт не может определить ваше имя электронной почты, пока вы не введете его. Разрешение веб-сайту создавать файлы cookie не дает этому или любому другому сайту доступа к остальной части вашего компьютера, и только сайт, который создал файл cookie, может его прочитать.
Развязка функций веретена-контрольной точки и хромосомной конгресса BubR1
Элоу, С., Дулла, К., Ульдшмид, А., Ли, X., Доу, З., и Нигг, Э.А. (2010) Разъединение функций веретено-контрольной точки и хромосомной конгресса BubR1. Журнал клеточной науки, Vol. 123, ч. 1. С. 84-94.
Полный текст недоступен в этом репозитории.
Официальный URL: http: //edoc.unibas.ch / dok / A5842440
Загрузки: Обзор статистики
Abstract
Белок контрольной точки BubR1 выполняет несколько функций в митозе. Мы провели функциональный анализ консервативных мотивов человеческого BubR1 (также известного как BUB1B) и продемонстрировали, что контрольная точка сборки веретена (SAC) и функции прикрепления хромосом могут быть отделены друг от друга. Мутация пяти пролин-направленных сайтов фосфорилирования серина, идентифицированных in vivo с помощью масс-спектрометрии, по существу отменяет прикрепление хромосом к веретену, но не влияет на функциональность SAC.Напротив, мутация двух консервативных боксов KEN, необходимых для функции SAC, не влияет на конгресс хромосом. Интересно, что вклад двух мотивов KEN-бокса неодинаков. Cdc20 связывается с N-концевым, но не с C-концевым KEN-боксом, а мутация N-концевого мотива KEN приводит к более сильному ускорению митотического времени. Более того, двух мотивов KEN недостаточно для максимального связывания Cdc20 и APC / C, что также требует последовательностей на С-конце BubR1. Наконец, мутация мотива GLEBS вызывает потерю взаимодействия Bub3 и неправильную локализацию BubR1 из кинетохоры; одновременно фосфорилирование BubR1, а также активность SAC и хромосомная конгрессия нарушаются, указывая на то, что мотив GLEBS строго необходим для обеих основных функций BubR1 человека.
| Факультеты и кафедры: | 05 Факультет естественных наук> Департамент Биоцентр> Бывшие организационные подразделения Биологический центр> Клеточная биология (Нигг) |
|---|---|
| UniBasel Участники: | Нигг, Эрих А. |