Хлоропласты | Биология
Хлоропласты являются одним из видов пластид. Хлоропласты имеют зеленый цвет за счет преобладающего в них пигмента хлорофилла. Основная их функция — фотосинтез.
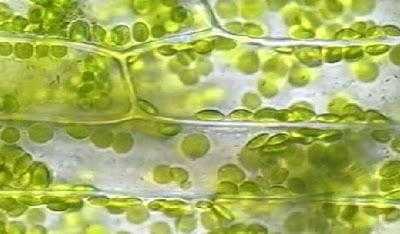
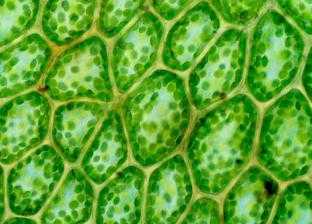
Количество данных органоидов в клетке варьирует. У некоторых водорослей в клетках содержится одни большой хлоропласт, часто причудливой формы. У высших растений их множество, особенно в мезофильной ткани листьев, где количество может достигать сотни штук на клетку.
У высших растений размер органоида около 5 мкм, форма округлая слегка вытянутая в одном направлении.

Хлоропласты в клетках развиваются из пропластид или путем деления надвое ранее существующих.
Строение хлоропласта
В строении хлоропластов выделяют внешнюю и внутреннюю мембраны, межмембранное пространство, строму, тилакоиды, граны, ламеллы, люмен.
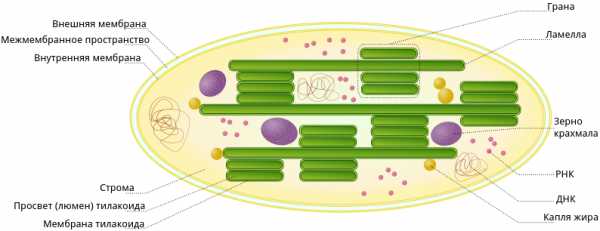
Тилакоид
Полужидкое содержимое хлоропласта называется стромой. В ней находятся его ДНК и РНК, рибосомы, обеспечивающие полуавтономность органоида (см. Симбиогенез).
Также в строме находятся зерна крахмала. Они образуются при избытке углеводов, образовавшихся при фотосинтетической активности. Жировые капли обычно формируются из мембран разрушающихся тилакоидов.
Функции хлоропластов
Основная функция хлоропластов — это фотосинтез — синтез глюкозы из углекислого газа и воды за счет солнечной энергии, которая улавливается хлорофиллом. В качестве побочного продукта фотосинтеза выделяется кислород. Однако процесс этот сложный и многоступенчатый, при котором синтезируются и побочные продукты, использующиеся как в самом хлоропласте, так и в остальных частях клетки.
Основным фотосинтетическим пигментом является хлорофилл. Он существует в нескольких разных формах. Кроме хлорофилла в фотосинтезе принимают участие пигменты каротиноиды.
Пигменты локализованы в мембранах тилакоидов, здесь протекают световые реакции фотосинтеза. Кроме пигментов здесь присутствуют ферменты и переносчики электронов. Хлоропласты стараются расположиться в клетке так, чтобы их тилакоидные мембраны находились под прямым углом к солнечному свету.
Хлорофилл состоит из длинного углеводного кольца и порфириновой головки. Хвост гидрофобен и погружен в липидный слой мембран тилакоидов. Головка гидрофильна и обращена к строме. Энергия света поглощается именно головкой, что приводит к возбуждению электронов.
Электрон отделяется от молекулы хлорофилла, который после этого становится электроположительным, т. е. оказывается в окисленной форме. Электрон принимается переносчиком, которые передает его на другое вещество.
Разные виды хлорофилла отличаются между собой несколько различным спектром поглощения солнечного света. Больше всего в растениях хлорофилла А.
В строме хлоропласта происходят темновые реакции фотосинтеза. Здесь находятся ферменты цикла Кальвина и другие.
biology.su
строение и функции в процессе фотосинтеза
Фотосинтез происходит в эукариотических клеточных структурах, называемых хлоропластами. Хлоропласт — это тип органеллы растительных клеток, известный как зеленые пластиды. Пластиды помогают хранить и собирать необходимые вещества для производства энергии. Хлоропласт содержит зеленый пигмент, называемый хлорофиллом, который поглощает световую энергию для процесса фотосинтеза. Следовательно, название хлоропласт указывает на то, что эти органеллы представляют собой хлорофиллсодержащие пластиды.
Подобно митохондриям, хлоропласты имеют свою собственную ДНК, ответственны за производство энергии и воспроизводятся независимо от остальной части клетки посредством процесса деления, подобного бактериальному бинарному делению. Они также ответственны за производство аминокислот и липидных компонентов, необходимых для производства хлоропластов. Хлоропласты также встречаются в клетках других фотосинтезирующих организмах, таких как водоросли.
Хлоропласт: структура
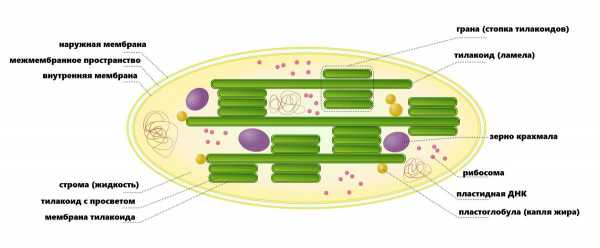
Схема строения хлоропласт
Хлоропласты обычно встречаются в охранных клетках, расположенных в листьях растений. Охранные клетки окружают крошечные поры, называемые устьицами, открывая и закрывая их, чтобы обеспечить необходимый для фотосинтеза газообмен. Хлоропласты и другие пластиды развиваются из клеток, называемых пропластидами, которые являются незрелыми, недифференцированными клетками, развивающимися в разные типы пластид. Пропластид, развивающийся в хлоропласт, осуществляет этот процесс только при свете. Хлоропласты содержат несколько различных структур, каждая из которых имеет специализированные функции. Основные структуры хлоропласта включают:
- Мембрана — содержит внутренние и внешние липидные двухслойные оболочки, которые выступают в качестве защитных покрытий и сохраняют замкнутые структуры хлоропластов. Внутренняя мембрана отделяет строму от межмембранного пространства и регулирует прохождение молекул в/из хлоропласта.
- Межмембранное пространство — пространство между внешней и внутренней мембранами.
- Тилакоидная система — внутренняя система мембран, состоящая из сплющенных мешкообразных мембранных структур, называемых тилакоидами, которые служат местами преобразования энергии света в химическую энергию.
- Тилакоид с просветом (люменом) — отсек в каждом тилакоиде.
- Грана — плотные слоистые стопки тилакоидных мешков (10-20), которые служат местами преобразования энергии света в химическую энергию.
- Строма — плотная жидкость внутри хлоропласта, содержащая внутри оболочки, но вне тилакоидной мембраны. Здесь происходит конверсия углекислого газа в углеводы (сахара).
- Хлорофилл — зеленый фотосинтетический пигмент в хлоропласт-гране, поглощающий световую энергию.
Хлоропласт: фотосинтез

При фотосинтезе энергия солнечного света преобразуется в химическую энергию. Химическая энергия хранится в виде глюкозы (сахара). Двуокись углерода, вода и солнечный свет используются для производства глюкозы, кислорода и воды. Фотосинтез происходит в два этапа: световая фаза и темновая фаза.
Световая фаза фотосинтеза протекает только при наличии света и происходит внутри хлоропластовой граны. Первичным пигментом, используемым для преобразования световой энергии в химическую, является хлорофилл а. Другие пигменты, участвующие в поглощении света, включают хлорофилл b, ксантофилл и каротин. Во время световой фазы, солнечный свет преобразуется в химическую энергию в виде АТФ (молекулы, содержащей свободную энергию) и НАДФ (молекула, несущая электроны высокой энергии).
И АТФ, и НАДФ используются во время темновой фазы для получения сахара. Темновая фаза фотосинтеза, также известная как этап фиксации углерода или цикл Кальвина. Реакции на этой стадии возникают в строме. Строма содержит ферменты, которые облегчают серию реакций, использующих АТФ, НАДФ и углекислый газ для получения сахара. Сахар может храниться в виде крахмала, используемого во время дыхания или при производстве целлюлозы.
Понравилась статья? Поделись с друзьями:
natworld.info
Хлоропласты, их состав, строение, свойства и функции
фотосинтез
протекает в специализированных органеллах
клеток — хлоропластах. Хлоропласты
высших растений имеют форму двояковыпуклой линзы (диска), которая наиболее удобна для
поглощения солнечных лучей. Их размеры,
количество, расположение полностью
отвечают назначению: как можно эффективнее
поглощать солнечную энергию, как можно
полнее усваивать углерод. Установлено,
что количество хлоропластов в клетке
измеряется десятками. Это обеспечивает
высокое содержание этих органелл на
единицу поверхности листа. Так, на
Строение хлоропласта весьма сложное. Подобно ядру и митохондриям хлоропласт окружен оболочкой, состоящей из двух липопротеидных мембран. Внутреннюю среду представляет относительно однородная субстанция — матрикс или
Свойства
хлоропластов: способны
измененять ориентацию и перемещаться. Например, под влиянием
яркого света хлоропласты поворачиваются
узкой стороной диска к падающим лучам
и перемещаются на боковые стенки клеток.
Хлоропласты передвигаются в направлении
более высокой концентрации СО
Химический состав хлоропластов: воды — 75 %; 75-80 % общего количества сухих веществ составляют орг. соединения, 20-25 % -минеральные.
Структурной основой хлоропластов являются белки (50-55 % сухой массы), половина из них составляют водорастворимые белки. Такое высокое содержание белков объясняется их многообразными функциями в составе хлоропластов (структурные белки мембран, белки-ферменты, транспортные белки, сократительные белки, реценторные).
Важнейшей составной частью хлоропластов являются липиды, (30-40% сух. м.). Липиды хлоропластов представлены тремя группами соединений.
Структурные компоненты мембран, которые представлены амфипатическими липоидами и отличаются высоким содержанием (более 50%) галактолипидов и сульфолипидов. Фосфолипидный состав характеризуется отсутствием фосфатидилэтаноламина и высоким содержанием фосфатидилглицерина (более 20 %). Свыше 60 % состава ЖК приходится на линолевую кислоту.
Фотосинтетическне пигменты хлоропластов — гидрофобные вв-а, относящиеся к липоидам (в клеточном соке — водорастворимые пигменты). Высшие растения содержат 2 формы зеленых пигментов: хлорофилл а и хлорофилл b и 2 формы желтых пигментов: каротины и ксантофиллы (каротиноиды). Хлорофиллы выполняет роль фотосенсибилизаторов, другие пигменты расширяют спектр действия фотосинтеза за счет более полного поглощения ФАР. Каротиноиды защищают хлорофилл от фотоокисления, участвуют в транспорте водорода, образующегося при фотолизе воды.
Жирорастворимые витамины — эргостерол (провитамин Д), витамины Е, К — сосредоточены практически целиком в хлоропластах, где участвуют в преобразовании световой энергии в химическую. В цитозоле клеток листа в основном находятся водорастворимые витамины. Так, у шпината содержание аскорбиновой кислоты в хлоропластах в 4-5 раз меньше, чем в листьях.
В хлоропластах листьев присутствует значительное количество РНК и ДНК. НК составляют примерно 1 % сухой массы хлоропластов (РНК — 0.75 %, ДНК — 0,01-0,02 %). Геном хлоропластов представлен кольцевой молекулой ДНК длиной 40 мкм с молекулярной массой 108, кодирующей 100-150 белков средних размеров. Рибосомы хлоропластов составляют от 20 до 50 % общей популяции рибосом клетки. Т.о., хлоропласты имеют собственную белоксинтезирующую систему. Однако для нормального функционирования хлоропластов необходимо взаимодеЯствие ядерного и хлоропластного геномов. Ключевой фермент фотосинтеза РДФ-карбоксилаза синтезируется под двойным контролем-ДНК ядра и хлоропласта.
Углеводы не являются конституционными веществами хлоропласта. Представлены фосфорными эфирами сахаров и продуктами фотосинтеза. Поэтому содержание углеводов в хлоропластах значительно колеблется (от 5 до 50 %). В активно функционирующих хлоропластах углеводы обычно не накапливаются, происходит их быстрый отток. При уменьшении потребности в продуктах фотосинтеза в хлоропластах образуются крупные крахмальные зерна. В этом случае содержание крахмала может возрасти до 50 % сухой массы и активность хлоропластов снизится.
Минеральные вещества. Сами хлоропласты составляют 25-30 % массы листа, но в них сосредоточено до 80 % Fe, 70-72 — Mg и Zn, 50 — Cu, 60 % Ca, содержащихся в тканях листа. Это объясняется высокой и разнообразной ферментативной активностью хлоропластов (входят с состав простетических групп и кофакторов). Mg входит в состав хлорофилла. Ca стабилизирует мембранные структуры хлоропластов.
Возникновение и развитие хлоропластов. Хлоропласты образуются в меристематических клетках из инициальных частиц или зачаточных пластид (рис.). Инициальная частица состоит из амебоидной стремы, окруженной двухмембранной оболочкой. По мере роста клетки инициалььные частицы увеличиваются в размере и приобретают форму двояковыпуклой линзы, в стреме появляются небольшие крахмальные зерна. Одновременно внутренняя мембрана начинает разрастаться, образуя складки (впячивания), от которых отшнуровываются пузырьки и трубочки. Такие образования называют пропластидами. Для дальнейшего их развития необходим свет. В темноте же формируются этиопласты, в которых образуется мембранная решетчатая структура — проламеллярное тело. На свету внутренние мембраны пропластид и этиопластов образуют гранильную систему. Одновременно с этим также на свету в граны встраиваются вновь образованные молекулы хлорофилла и других пигментов. Таким образом, структуры, которые подготавливаются к функционированию на свету, появляются и развиваются только при его наличии.
Наряду с хлоропластами имеется ряд других пластид, которые образуются либо непосредственно из пропластид, либо одна из другой путем взаимных превращений (рис.). К ним относятся накапливающие крахмал амилопласты (лейкопласты) и хромопласты, содержащие каротиноиды. В цветках и плодах хромопласты возникают на ранних стадиях развития пропластид. Хромопласты осенней листвы представляют собой продукты деградации хлоропластов, в которых в качестве структур — носителей каротнноидов выступают пластоглобулы.
Пигменты хлоропласта, участвующие в улавливании световой энергии, а также ферменты, необходимые для световой фазы фотосинтеза, вмонтированы в мембраны тилакоидов.
Ферменты, которые катализируют многочисленные реакции восстановительного цикла углеводов (темповой фазы фотосинтеза), а также разнообразные биосинтезы, в том числе биосинтезы белков, липидов, крахмала, присутствуют главным образом в строме, часть из них является периферическими белками ламелл.
Строение зрелых хлоропластов одинаково у всех высших растений, так же как в клетках разных органов одного растения (листьях, зеленеющих корнях, коре, плодах). В зависимости от функциональной нагрузки клеток, физиологического состояния хлоропластов, их возраста различают степень их внутренней структурированности: размеры, количество гран, связь между ними. Так, в замыкающих клетках устьиц основная функция хлоропластов — фоторегуляция устьичных движений. Хлоропласты не имеют строгой гранальной структуры, содержат крупные крахмальные зерна, набухшие тилакоиды, липофильные глобулы. Все это свидетельствует об их низкой энергетической нагрузке (эту функцию выполняют митохондрии). Другая картина наблюдается при изучении хлоропластов зеленых плодов томата. Наличие хорошо развитой гранулярной системы свидетельствует о высокой фукциональной нагрузке этих органелл и, вероятно, существенном вкладе фотосинтеза при формировании плодов.
Возрастные изменения: Молодые характеризуются ламеллярнои структурой, в таком состоянии хлоропласты способны размножаться путем деления. В зрелых хорошо выражена система гран. В стареющих происходит разрыв тилакоидов стромы, связь между гранами уменьшается, в дальнейшем наблюдаются распад хлорофилла и деструкция гран. В осенней листве деградация хлоропластов приводит к образованию хромопластов.
Структура хлоропластов лабильна и динамична, в ней отражаются все условия жизни растения. Большое влияние оказывает режим минерального питания растений. При недостатке N хлоропласты становятся в 1.5-2 раза мельче, дефицит P и S нарушает нормальную структуру ламелл и гран, одновременная нехватка N и Ca приводит к переполнению хлоропластов крахмалом из-за нарушения нормального оттока ассимилятов. При недостатке Ca нарушается структура наружной мембраны хлоропласта. Для поддержания структуры хлоропласта также необходим свет, в темноте идет постепенное разрушение тилакоидов гран и стремы.
studfile.net
Внутренняя мембрана хлоропластов — Справочник химика 21
ТОННОГО градиента. Интактные клетки реагируют на воздействие света выделением в среду протонов, приводящим к закислению среды. В суспензии пузырьков из фотосинтетических мембран (хроматофоров) свет вызывает перенос протонов, направленный внутрь. Таким образом, мембраны хроматофоров и тилакоидов имеют такую же полярность, как и субмитохондриальные пузырьки. Это будет поняты , если учесть, что все эти мембраны образуются путем впячивания внутрь и разрастания плазматической мембраны или же внутренней мембраны хлоропласта. Хотя точная локализация отдельных компонентов в мембране еще не установлена, можно думать, что переносчики водорода и электронов расположены и в мембране анаэробных фототрофных бактерий таким образом, что происходит разделение зарядов. В хроматофорах электроны транспортируются наружу, а протоны-внутрь. Создающийся протонный потенциал и служит движущей силой фотосинтетического фосфорилирования. [c.392]Внутренняя мембрана хлоропластов образует уплошенные мембранные тела — пузырьковидные диски, называемые тилакоидными дисками, которые уложены в стопку и соединены между собой перемычками. Эта стопка называется граной, их число составляет 40—80 шт. на один хлоропласт. Наружная мембрана хлоропластов, как и митохондрий, гладкая (рис. 16.1). [c.211]
A. Внутренняя мембрана хлоропласта окружает большую центральную область, называемую , которая представляет собой аналог митохондриального матрикса. [c.84]
Однако есть и существенное отличие Внутренняя мембрана хлоропластов не образует крист и не содержит в себе цепи переноса электронов. Фотосинтезирующая поглощающая свет система, электрон-транспортная цепь и АТР-синтетаза находятся в третьей мембране. [c.461]
Внутренняя мембрана хлоропласта, подобно внутренней мембране митохондрии, содержит белки-переносчики, 7.5.12. облегчающие обмен метаболитами с цитозолем 476 [c.515]
Хлоропласты зеленых растений, в которых находится пигмент хлорофилл, имеют весьма сложное строение. Они одеты двойной мембраной. В их содержимом — строме — имеется хорошо развитая система мембранных дисков (тилакоидов), которые могут быть собраны в стопки, именуемые гранами. На мембранах тилакоидов или в этих мембранах локализованы пигменты, участвующие в процессе фотосинтеза, переносчики электронов (цитохромы, пластоцианин, пластохинон, ферредо-ксин) и фермент АТРаза таким образом, здесь могут протекать процессы, характерные для фотосинтеза, — зависящие от света окислительно-восстановительные реакции и синтез АТР. Фиксация СО2 и ее превращение в сахара в результате биосинтетических реакций протекают в строме, где находится соответствующий набор ферментов. Внутренняя мембрана хлоропластов имеет ограниченную проницаемость для метаболитов и, подобно внутренней мембране митохондрий, снабжена специфической транспортной системой, регулирующей перенос веществ внутрь Стромы и из нее. [c.93]
Однако есть и существенное отличие. Внутренняя мембрана хлоропластов не образует крист и не содержит в себе цепи переноса электронов. Фотосинтезирующая поглощающая свет система, электронтранспортная цепь и АТР-синтетаза находятся в третьей мембране, формирующей группу уплощенных дисковидных мешочков-тилакоидов (рис. 9-37). Внутренние полости тилакои-дов сообщаются между собой, образуя третий внутренний компартмент хлоропласта, называемый тилакоидным пространством. Тилакоидное пространство отделено от стромы непроницаемой для ионов тилакоидной мембраной. [c.36]
Д. Модель, суммирующая роль сигнальных пептидов в импорте ферредоксина и пластоцианина в соответствующие компартменты хлоропластов, изображена на рис. 8-22. Для каждого белка сигнальный пептид показан как прямоугольник на N-конце белка. Зачерненная часть прямоугольника соответствует той части сигнального пептида, которая необходима для импорта через наружную и внутреннюю мембраны хлоропласта. Этот сигнальный пептид отделяется, когда белок пересекает мембрану и входит в строму. Незачерненная часть прямоугольника, связанная с пластоцианином, нужна для импорта через тилакоидную мембрану. Она удаляется при импорте в полость тилакоида. [c.370]
| Рис. 3.6. Хлоропласты бобов в забуференном растворе сахарозы. Внутренние мембраны хлоропластов интактны, но наружной оболочки нет. По классификации, приведенной в гл. 3, хлоропласты принадлежат к типу С. [Печатается с любезного разрешения А. Гринвуда и Р. Лича (Greenwood, Lee h).] |
www.chem21.info
Мембраны хлоропластов — Справочник химика 21
Прокариотная клетка отличается тем, что имеет одну внутреннюю полость, образуемую элементарной мембраной, называемой клеточной, или цитоплазматической (ЦПМ). У подавляющего большинства прокариот ЦПМ — единственная мембрана, обнаруживаемая в клетке. В эукариотных клетках в отличие от прокариотных есть вторичные полости. Ядерная мембрана, отграничивающая ДНК от остальной цитоплазмы, формирует вторичную полость. Наружные мембраны хлоропластов и митохондрий, окружающие заключенные в них функционально специализированные мембраны, играют аналогичную роль. Клеточные структуры, Офаниченные элементарными мембранами и выполняющие в клетке определенные функции, получили название органелл. Ядро, митохондрии, хлоропласты — это клеточные органеллы. В эукариотных клетках помимо перечисленных выше есть и другие органеллы. [c.18]
Ультраструктура тилакоидной мембраны хлоропласта [c.332]
ТОННОГО градиента. Интактные клетки реагируют на воздействие света выделением в среду протонов, приводящим к закислению среды. В суспензии пузырьков из фотосинтетических мембран (хроматофоров) свет вызывает перенос протонов, направленный внутрь. Таким образом, мембраны хроматофоров и тилакоидов имеют такую же полярность, как и субмитохондриальные пузырьки. Это будет поняты , если учесть, что все эти мембраны образуются путем впячивания внутрь и разрастания плазматической мембраны или же внутренней мембраны хлоропласта. Хотя точная локализация отдельных компонентов в мембране еще не установлена, можно думать, что переносчики водорода и электронов расположены и в мембране анаэробных фототрофных бактерий таким образом, что происходит разделение зарядов. В хроматофорах электроны транспортируются наружу, а протоны-внутрь. Создающийся протонный потенциал и служит движущей силой фотосинтетического фосфорилирования. [c.392]
Хлоропласты представляют интерес в первую очередь в связи с их фотосинтетической функцией. Однако с более общей точки зрения фотосинтез — это лишь одна из многих важных биологических функций хлоропластов. Недавно было показано, что хлоропласты способны синтезировать белок, что опи изменяют свою форму и подвергаются конформационным изменениям в процессе переноса электронов [25] и что мембрана хлоропластов способна к накоплению ионов [30]. Генетическая автономность хлоропластов, связанная с наличием в них нуклеиновых кислот, способы самовоспроизведения хлоропластов и их метаболические функции, отличные от фотосинтеза, — вот интереснейшие вопросы, ждущие своего разрешения. Здесь будут рассмотрены некоторые из этих проблем. [c.81]
Мембраны хлоропластов 0,6 Фотосинтез [c.106]
Еще один подход, позволяющий судить о транспортных продуктах фотосинтеза — изучение оттока различных соединений из изолированных хлоропластов в окружающую водную среду. Естественно, что в этом случае необходимым требованием является целостность внешней мембраны хлоропластов. Осуществление кинетических опытов позволяет судить о наиболее быстрых транспортных продуктах и об их превращении в другие соединения вне хлоропластов. [c.260]
Биоэнергетические процессы, приводящие к синтезу АТФ, к зарядке биологических аккумуляторов , протекают в мембранах митохондрий. В них локализованы и пространственно организованы молекулярные системы, ответственные за энергетику живых организмов. Синтез АТФ в митохондриях сопряжен с электронным и ионным транспортом и с механохимическими явлениями. Функции митохондриальных мембран весьма сложны и многообразны. Другой тип биоэнергетических сопрягающих мембран — мембраны хлоропластов растений, ответственные за фотосинтез,— рассматривается в гл. 14. У бактерий сопряжение реализуется в плазматических мембранах. [c.423]
Внутренняя мембрана хлоропластов образует уплошенные мембранные тела — пузырьковидные диски, называемые тилакоидными дисками, которые уложены в стоп
www.chem21.info
Единицы живого: Хлоропласты
Хлоропласты – это структуры, в которых происходят фотосинтетические процессы, приводящие в конечном итоге к связыванию углекислоты, к выделению кислорода и синтезу сахаров. структуры удлиненной формы с шириной 2-4 мкм и протяженностью 5-10 мкм. У зеленых водорослей встречаются гигантские хлоропласты (хроматофоры), достигающие длины 50 мкм.
у зеленых водорослей может быть по одному хлоропласту на клетку. Обычно на клетку высших растений приходится в среднем 10-30 хлоропластов. Встречаются клетки с огромным количеством хлоропластов. Например, в гигантских клетках палисадной ткани махорки обнаружено около 1000 хлоропластов.
Хлоропласты представляют собой структуры, ограниченные двумя мембранами – внутренней и внешней. Внешняя мембрана, как и внутренняя, имеет толщину около 7 мкм, они отделены друг от друга межмембранным пространством около 20-30 нм. Внутренняя мембрана хлоропластов отделяет строму пластиды, аналогичную матриксу митохондрий. В строме зрелого хлоропласта высших растений видны два типа внутренних мембран. Это – мембраны, образующие плоские, протяженные ламеллы стромы, и мембраны тилакоидов, плоских дисковидных вакуолей или мешков.
Ламеллы стромы (толщиной около 20 мкм) представляют собой плоские полые мешки или же имеют вид сети из разветвленных и связанных друг с другом каналов, располагающихся в одной плоскости. Обычно ламеллы стромы внутри хлоропласта лежат параллельно друг другу и не образуют связей между собой.
Кроме мембран стромы в хлоропластах обнаруживаются мембранные тилакоиды. Это плоские замкнутые мембранные мешки, имеющие форму диска. Величина межмембранного пространства у них также около 20-30 нм. Такие тилакоиды образуют стопки наподобие столбика монет, называемые гранами.
Число тилакоидов на одну грану очень варьирует: от нескольких штук до 50 и более. Размер таких стопок может достигать 0,5 мкм, поэтому граны видны в некоторых объектах в световом микроскопе. Количество гран в хлоропластах высших растений может достигать 40-60. Тилакоиды в гране сближены друг с другом так, что внешние слои их мембран тесно соединяются; в месте соединения мембран тилакоидов образуется плотный слой толщиной около 2 нм. В состав граны кроме замкнутых камер тилакоидов обычно входят и участки ламелл, которые в местах контакта их мембран с мембранами тилакоидов тоже образуют плотные 2-нм слои. Ламеллы стромы, таким образом, как бы связывают между собой отдельные граны хлоропласта. Однако полости камер тилакоидов всезда замкнуты и не переходят в камеры межмембранного пространства ламелл стромы. Ламеллы стромы и мембраны тилакоидов образуются путем отделения от внутренней мембраны при начальных этапах развития пластид.
В матриксе (строме) хлоропластов обнаруживаются молекулы ДНК, рибосомы; там же происходит первичное отложение запасного полисахарида, крахмала, в виде крахмальных зерен.
Характерным для хлоропластов является наличие в них пигментов, хлорофиллов, которые и придают окраску зеленым растениям. При помощи хлорофилла зеленые растения поглощают энергию солнечного света и превращают ее в химическую.
Функции хлоропластов
Геном пластид
Подобно митохондриям, хлоропласты имеют собственную генетическую систему, обеспечивающую синтез ряда белков внутри самих пластид. В матриксе хлоропластов обнаруживаются ДНК, разные РНК и рибосомы. Оказалось, что ДНК хлоропластов резко отличается от ДНК ядра. Она представлена циклическими молекулами длиной до 40-60 мкм, имеющими молекулярный вес 0,8-1,3х108 дальтон. В одном хлоропласте может быть множество копий ДНК. Так, в индивидуальном хлоропласте кукурузы присутствует 20-40 копий молекул ДНК. Длительность цикла и скорость репликации ядерной и хлоропластной ДНК, как было показано на клетках зеленых водорослей, не совпадают. ДНК хлоропластов не состоит в комплексе с гистонами. Все эти характеристики ДНК хлоропластов близки к характеристикам ДНК прокариотических клеток. Более того, сходство ДНК хлоропластов и бактерий подкрепляется еще и тем, что основные регуляторные последовательности транскрипции (промоторы, терминаторы) у них одинаковы. На ДНК хлоропластов синтезируются все виды РНК (информационная, трансферная, рибосомная). ДНК хлоропластов кодирует рРНК, входящую в состав рибосом этих пластид, которые относятся к прокариотическому 70S типу (содержат 16S и 23S рРНК). Рибосомы хлоропластов чувствительны к антибиотику хлорамфениколу, подавляющему синтез белка у прокариотических клеток.
Так же как в случае хлоропластов мы вновь сталкиваемся с существованием особой системы синтеза белка, отличной от таковой в клетке.
Эти открытия вновь пробудили интерес к теории симбиотического происхождения хлоропластов. Идея о том, что хлоропласты возникли за счет объединения клеток-гетеротрофов с прокариотическими синезелеными водорослями, высказанная на рубеже XIX и XX вв. (А.С. Фоминцин, К.С.Мережковский) вновь находит свое подтверждение. В пользу этой теории говорит удивительное сходство в строении хлоропластов и синезеленых водорослей, сходство с основными их функциональными особенностями, и в первую очередь со способностью к фотосинтетическим процессам.
Известны многочисленные факты истинного эндосимбиоза синезеленых водорослей с клетками низших растений и простейших, где они функционируют и снабжают клетку-хозяина продуктами фотосинтеза. Оказалось, что выделенные хлоропласты могут также отбираться некоторыми клетками и использоваться ими как эндосимбионты. У многих беспозвоночных (коловратки, моллюски), питающихся высшими водорослями, которые они переваривают, интактные хлоропласты оказываются внутри клеток пищеварительных желез. Так, у некоторых растительноядных моллюсков в клетках найдены интактные хлоропласты с функционирующими фотосинтетическими системами, за активностью которых следили по включению С14О2.
Как оказалось, хлоропласты могут быть введены в цитоплазму клеток культуры фибробластов мыши путем пиноцитоза. Однако они не подвергались атаке гидролаз. Такие клетки, включившие зеленые хлоропласты, могли делиться в течение пяти генераций, а хлоропласты при этом оставались интактными и проводили фотосинтетические реакции. Были предприняты попытки культивировать хлоропласты в искусственных средах: хлоропласты могли фотосинтезировать, в них шел синтез РНК, они оставались интактными 100 ч, у них даже в течение 24 ч наблюдались деления. Но затем происходило падение активности хлоропластов, и они погибали.
Эти наблюдения и целый ряд биохимических работ показали, что те черты автономии, которыми обладают хлоропласты, еще недостаточны для длительного поддержания их функций и тем более для их воспроизведения.
В последнее время удалось полностью расшифровать всю последовательность нуклеотидов в составе циклической молекулы ДНК хлоропластов высших растений. Эта ДНК может кодировать до 120 генов, среди них: гены 4 рибосомных РНК, 20 рибосомных белков хлоропластов, гены некоторых субъединиц РНК-полимеразы хлоропластов, несколько белков I и II фотосистем, 9 из 12 субъединиц АТФ-синтетазы, части белков комплексов цепи переноса электронов, одной из субъединиц рибулозодифосфат-карбоксилазы (ключевой фермент связывания СО2), 30 молекул тРНК и еще 40 пока неизвестных белков. Интересно, что сходный набор генов в ДНК хлоропластов обнаружен у таких далеко отстоящих представителей высших растений как табак и печеночный мох.
Основная же масса белков хлоропластов контролируется ядерным геномом. Оказалось, что ряд важнейших белков, ферментов, а соответственно и метаболические процессы хлоропластов находятся под генетическим контролем ядра. Так, клеточное ядро контролирует отдельные этапы синтеза хлорофилла, каротиноидов, липидов, крахмала. Под ядерным контролем находятся многие энзимы темновой стадии фотосинтеза и другие ферменты, в том числе некоторые компоненты цепи транспорта электронов. Ядерные гены кодируют ДНК-полимеразу и аминоацил-тРНК-синтетазу хлоропластов. Под контролем ядерных генов находится большая часть рибосомных белков. Все эти данные заставляют говорить о хлоропластах, так же как и о митохондриях, как о структурах с ограниченной автономией.
Транспорт белков из цитоплазмы в пластиды происходит в принципе сходно с таковым у митохондрий. Здесь также в местах сближения внешней и внутренней мембран хлоропласта располагаются каналообразующие интегральные белки, которые узнают сигнальные последовательности хлоропластных белков, синтезированных в цитоплазме, и транспортируют их в матрикс-строму. Из стромы импортируемые белки согласно дополнительным сигнальным последовательностям могут включаться в мембраны пластиды (тилакоиды, ламеллы стромы, внешняя и внутренняя мембраны) или локализоваться в строме, входя в состав рибосом, ферментных комплексов цикла Кальвина и др.
Удивительное сходство структуры и энергетических процессов у бактерий и митохондрий, с одной стороны, и у синезеленых водорослей и хлоропластов – с другой, служит веским аргументом в пользу теории симбиотического происхождения этих органелл. Согласно этой теории, возникновение эукариотической клетки прошло через несколько этапов симбиоза с другими клетками. На первой стадии клетки типа анаэробных гетеротрофных бактерий включили в себя аэробные бактерии, превратившиеся в митохондрии. Параллельно этому в клетке-хозяине прокариотический генофор формируется в обособленное от цитоплазмы ядро. Так могли возникнуть гетеротрофные эукариотические клетки. Повторные эндосимбиотические взаимоотношения между первичными эукариотическими клетками и синезелеными водорослями привели к появлению в них структур типа хлоропластов, позволяющих клеткам осуществлять автосинтетические процессы и не зависеть от наличия органических субстратов (рис. 236). В процессе становления такой составной живой системы часть генетической информации митохондрий и пластид могла изменяться, перенестись в ядро. Так, например две трети из 60 рибосомных белков хлоропластов кодируется в ядре и синтезируются в цитоплазме, а потом встраивается в рибосомы хлоропластов, имеющие все свойства прокариотических рибосом. Такое перемещение большой части прокариотических генов в ядро привело к тому, что эти клеточные органеллы, сохранив часть былой автономии, попали под контроль клеточного ядра, определяющего в большей степени все главные клеточные функции.
Пропластиды
При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов. Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов.
При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы (рис. 226 б). Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО2, то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов, сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды (рис. 226в).
Другой формой пластид у высших растений является хромопласт, окрашивающийся обычно в желтый свет в результате накопления в нем каротиноидов (рис. 226г). Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы сопряжены с постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.
biology623.blogspot.com
Что такое хлоропласт? Хлоропласты: строение и функции
Растительный мир — одно из главных богатств нашей планеты. Именно благодаря флоре на Земле есть кислород, которым мы все дышим, имеется огромная пищевая база, от которой зависит все живое. Растения уникальны тем, что могут превращать химические соединения неорганической природы в органические вещества.
 Делают они это посредством фотосинтеза. Этот важнейший процесс протекает в специфических растительных органоидах, хлоропластах. Этот мельчайший элемент фактически обеспечивает существование всей жизни на планете. Кстати, а что такое хлоропласт?
Делают они это посредством фотосинтеза. Этот важнейший процесс протекает в специфических растительных органоидах, хлоропластах. Этот мельчайший элемент фактически обеспечивает существование всей жизни на планете. Кстати, а что такое хлоропласт?Основное определение
Так называются специфические структуры, в которых происходят процессы фотосинтеза, которые направлены на связывание углекислого газа и образование некоторых углеводов. Побочным продуктом является кислород. Это вытянутые в длину органоиды, достигающие в ширину 2-4 мкм, длина их доходит до 5-10 мкм. У некоторых видов зеленых водорослей порой встречаются хлоропласты-гиганты, вытянутые на 50 мкм!
У этих же водорослей может быть другая особенность: на всю клетку у них имеется только один органоид этого вида. В клетках высших растений чаще всего имеется в пределах 10-30 хлоропластов. Впрочем, и в их случае могут встречаться яркие исключения. Так, в палисадной ткани обычной махорки имеется по 1000 хлоропластов на одну клетку. Для чего нужны эти хлоропласты? Фотосинтез – вот их главная, но далеко не единственная роль. Чтобы четко понимать их значение в жизни растения, важно знать многие аспекты их происхождения и развития. Все это описывается в дальнейшей части статьи.
Происхождение хлоропласта
Итак, что такое хлоропласт, мы узнали. А откуда эти органоиды произошли? Как получилось, что у растений появился столь уникальный аппарат, который превращает углекислый газ и воду в сложные органические соединения?
В настоящее время среди ученых превалирует точка зрения об эндосимбиотическом происхождении данных органоидов, так как их самостоятельное возникновение в клетках растения довольно сомнительно. Отлично известно, что лишайник – это симбиоз водоросли и гриба. Одноклеточные водоросли при этом живут внутри грибной клетки. Сейчас ученые предполагают, что в незапамятные времена фотосинтезирующие цианобактерии проникли внутрь растительных клеток, а затем частично утратили «самостоятельность», передав большую часть генома в ядро.
Но свою главную особенность новый органоид сохранил в полной мере. Речь идет как раз о процессе фотосинтеза. Впрочем, сам аппарат, необходимый для выполнения данного процесса, формируется под контролем как клеточного ядра, так и самого хлоропласта. Так, деление этих органоидов и прочие процессы, связанные с реализацией генетической информации на ДНК, контролируются ядром.Доказательства
Относительно недавно гипотеза о прокариотическом происхождении этих элементов была не слишком популярна в научном сообществе, многие считали ее «измышлениями дилетантов». Но после того как был проведен углубленный анализ нуклеотидных последовательностей в ДНК хлоропластов, это предположение получило блестящее подтверждение. Выяснилось, что эти структуры чрезвычайно схожи, даже родственны, ДНК бактериальных клеток. Так, аналогичная последовательность была найдена у свободноживущих цианобактерий. В частности, оказались чрезвычайно схожи гены АТФ-синтезирующего комплекса, а также в «аппаратах» транскрипции и трансляции.
Промоторы, которые определяют начало считывания генетической информации с ДНК, а также терминальные нуклеотидные последовательности, которые отвечают за ее прекращение, также организованы по образу и подобию бактериальных. Разумеется, миллиарды лет эволюционных преобразований смогли внести множество изменений в хлоропласт, но последовательности в хлоропластных генах остались абсолютно прежними. И это – неопровержимое, полное доказательство того, что хлоропласты и в самом деле когда-то имели прокариотического предка. Возможно, это был организм, от которого произошли также современные цианобактерии.
Развитие хлоропласта из пропластиды
«Взрослый» органоид развивается из пропластиды. Это маленькая, полностью бесцветная органелла, имеющая всего несколько микрон в поперечнике. Она окружена плотной двуслойной мембраной, которая содержит кольцевую ДНК, специфическую для хлоропласта. Внутренней мембранной системы эти «предки» органоидов не имеют. Из-за предельно малых размеров их изучение крайне затруднено, а потому данных об их развитии чрезвычайно мало.
Известно, что несколько таких протопластид имеется в ядре каждой яйцеклетки животных и растений. В ходе развития зародыша они делятся и передаются другим клеткам. Это легко проверить: генетические признаки, которые так или иначе связаны с пластидами, передаются только по материнской линии.
Внутренняя мембрана протопластиды за время развития выпячивается внутрь органоида. Из этих структур вырастают мембраны тилакоидов, которые отвечают за образование гран и ламелл стромы органоида. В полной темноте протопастида начинает преобразовываться в предшественник хлоропласта (этиопласта). Этот первичный органоид характерен тем, что внутри него располагается довольно сложная кристаллическая структура. Как только на лист растения попадет свет, она полностью разрушается. После этого происходит образование «традиционной» внутренней структуры хлоропласта, которая образована как раз-таки тилакоидами и ламеллами.
Отличия растений, запасающих крахмал
В каждой меристемальной клетке содержится несколько таких пропластид (их количество разнится в зависимости от вида растения и прочих факторов). Как только эта первичная ткань начинает преобразовываться в лист, предшественники органоидов превращаются в хлоропласты. Так, закончившие свой рост молодые листья пшеницы имеют хлоропласты в количестве 100-150 штук. Чуть сложнее обстоят дела в отношении тех растений, которые способны к накоплению крахмала.
 Они скапливают запас этого углевода в пластидах, которые именуются амилопластами. Но какое отношение эти органоиды имеют к теме нашей статьи? Ведь клубни картофеля не участвуют в фотосинтезе! Позвольте разъяснить этот вопрос более подробно.
Они скапливают запас этого углевода в пластидах, которые именуются амилопластами. Но какое отношение эти органоиды имеют к теме нашей статьи? Ведь клубни картофеля не участвуют в фотосинтезе! Позвольте разъяснить этот вопрос более подробно.Мы выяснили, что такое хлоропласт, попутно выявив связь этого органоида со структурами прокариотических организмов. Здесь ситуация схожа: ученые давно выяснили, что амилопласты, как и хлоропласты, содержат точно такую же ДНК и образуются из точно тех же протопластид. Следовательно, и рассматривать их следует в том же аспекте. Фактически амилопласты следует рассматривать в качестве особой разновидности хлоропласта.
Как образуются амилопласты?
Можно провести аналогию между протопластидами и стволовыми клетками. Проще говоря, амилопласты с какого-то момента начинают развиваться по несколько иному пути. Ученые, впрочем, узнали кое-что любопытное: им удалось добиться взаимного превращения хлоропластов из листьев картофеля в амилопласты (и наоборот). Каноничный пример, известный каждому школьнику – клубни картофеля на свету зеленеют.
Прочие сведения о путях дифференцирования этих органоидов
Мы знаем, что в процессе созревания плодов томата, яблок и некоторых других растений (и в листьях деревьев, трав и кустарников в осенний период) происходит процесс «деградации», когда хлоропласты в растительной клетке превращаются в хромопласты. Эти органоиды содержат в своем составе красящие пигменты, каротиноиды.
Превращение это связано с тем, что в определенных условиях происходит полное разрушение тилакоидов, после чего органелла приобретает иную внутреннюю организацию. Вот здесь-то мы снова возвращаемся к тому вопросу, который начали обсуждать в самом начале статьи: влияние ядра на развитие хлоропластов. Именно оно, посредством особых белков, которые синтезируются в цитоплазме клеток, инициирует процесс перестройки органоида.
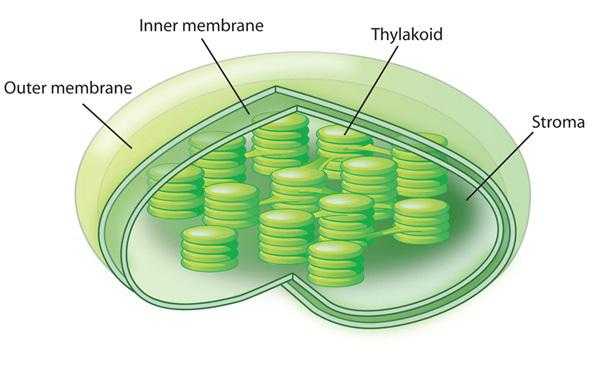
Строение хлоропласта
Поговорив о вопросах происхождения и развития хлоропластов, следует подробнее остановиться на их строении. Тем более что оно весьма интересно и заслуживает отдельного обсуждения.
Основная структура хлоропластов состоит из двух липопротеиновых мембран, внутренней и внешней. Толщина каждой составляет порядка 7 нм, расстояние между ними — 20-30 нм. Как и в случае других пластид, внутренний слой образует особые структуры, выпячивающиеся внутрь органоида. У зрелых хлоропластов существует сразу два типа таких «извилистых» мембран. Первые образуют ламеллы стромы, вторые – мембраны тилакоидов.
Ламеллы и тилакоиды
Нужно заметить, что прослеживается четкая связь, которую имеет мембрана хлоропластов с аналогичными образованиями, находящимися внутри органоида. Дело в том, что некоторые ее складки могут простираться от одной стенки до другой (как у митохондрий). Так что ламеллы могут образовывать либо своеобразный «мешок», либо разветвленную сеть. Впрочем, чаще всего эти структуры располагаются параллельно друг другу и никак не связаны между собой.
 Не стоит забывать, что внутри хлоропласта находятся еще и мембранные тилакоиды. Это замкнутые «мешки», которые располагаются в виде стопки. Как и в предыдущем случае, между двумя стенками полости имеется расстояние в 20-30 нм. Столбики из этих «мешков» называются гранами. В каждом столбике может находиться до 50 тилакоидов, а в некоторых случаях их бывает еще больше. Так как общие «габариты» таких стопок могут достигать 0,5 мкм, иногда они могут быть обнаружены при помощи обыкновенного светового микроскопа.
Не стоит забывать, что внутри хлоропласта находятся еще и мембранные тилакоиды. Это замкнутые «мешки», которые располагаются в виде стопки. Как и в предыдущем случае, между двумя стенками полости имеется расстояние в 20-30 нм. Столбики из этих «мешков» называются гранами. В каждом столбике может находиться до 50 тилакоидов, а в некоторых случаях их бывает еще больше. Так как общие «габариты» таких стопок могут достигать 0,5 мкм, иногда они могут быть обнаружены при помощи обыкновенного светового микроскопа.Общее количество гран, которые содержатся в хлоропластах высших растений, может доходить до 40-60. Каждый тилакоид так плотно прилегает к другому, что их внешние мембраны образуют единую плоскость. Толщина слоя в месте соединения может доходить до 2 нм. Заметим, что подобные структуры, которые образованы прилегающими друг к другу тилакоидами и ламеллами, совсем нередки.
В местах их соприкосновения также имеется слой, достигающий порой тех же самых 2 нм. Таким образом, хлоропласты (строение и функции которых весьма сложны) представляют собой не единую монолитную структуру, а своеобразное «государство внутри государства». В некоторых аспектах строение этих органоидов не менее сложно, чем вся клеточная структура!
Граны связываются между собой именно при помощи ламелл. Но полости тилакоидов, которые образуют стопки, всегда замкнуты и никак не сообщаются с межмембранным пространством. Как видите, структура хлоропластов достаточно сложна.
Какие пигменты могут содержаться в хлоропластах?
Что может содержаться в строме каждого хлоропласта? Там имеются отдельные молекулы ДНК и немало рибосом. У амилопластов именно в строме откладываются крахмальные зерна. Соответственно, у хромопластов там имеются красящие пигменты. Разумеется, встречаются различные пигменты хлоропластов, но наиболее распространенным является хлорофилл. Он подразделяется сразу на несколько видов:
- Группа А (сине-зеленый). Встречается в 70% случаев, содержится в хлоропластах всех высших растений и водорослей.
- Группа В (желто-зеленый). В остальных 30% также обнаруживается у растений и водорослей высших видов.
- Группы С, D и Е встречаются намного реже. Имеются в хлоропластах некоторых видов низших водорослей и растений.
У красных и бурых морских водорослей в хлоропластах не так уж и редко могут иметься совершенно другие виды органических красителей. В некоторых же водорослях вообще содержатся едва ли не все существующие пигменты хлоропластов.
Функции хлоропластов
Разумеется, основной их функцией является преобразование световой энергии в органические компоненты. Сам фотосинтез происходит в гранах при непосредственном участии хлорофилла. Он поглощает энергию солнечного света, переводя ее в энергию возбужденных электронов. Последние, обладая избыточным ее запасом, отдают излишки энергии, которая используется для разложения воды и синтеза АТФ. При распаде воды образуется кислород и водород. Первый, как мы уже писали выше, является побочным продуктом и выделяется в окружающее пространство, а водород связывается с особым белком, ферредоксином.
 Он снова окисляется, передавая водород восстановителю, который в биохимии обозначается аббревиатурой НАДФ. Соответственно, его восстановленная форма — НАДФ-h3. Проще говоря, в процессе фотосинтеза происходит выделение следующих веществ: АТФ, НАДФ-h3 и побочного продукта в виде кислорода.
Он снова окисляется, передавая водород восстановителю, который в биохимии обозначается аббревиатурой НАДФ. Соответственно, его восстановленная форма — НАДФ-h3. Проще говоря, в процессе фотосинтеза происходит выделение следующих веществ: АТФ, НАДФ-h3 и побочного продукта в виде кислорода.Энергетическая роль АТФ
Образующаяся АТФ крайне важна, так как является основным «аккумулятором» энергии, которая идет на различные нужды клетки. НАДФ-h3 содержит восстановитель, водород, причем это соединение способно легко его отдавать в случае необходимости. Проще говоря, это эффективный химический восстановитель: в процессе фотосинтеза происходит множество реакций, которые без него попросту не смогут протекать.
Далее в дело вступают ферменты хлоропластов, которые действуют в темноте и вне гран: водород из восстановителя и энергия АТФ используются хлоропластом для того, чтобы начать синтез ряда органических веществ. Так как фотосинтез происходит в условиях хорошей освещенности, накопленные соединения в темное время суток используются для нужд самих растений.
Вы справедливо можете заметить, что этот процесс в некоторых аспектах подозрительно похож на дыхание. Чем отличается от него фотосинтез? Таблица поможет вам разобраться в этом вопросе.
Пункты сравнения | Фотосинтез | Дыхание |
Когда происходит | Только днем, при солнечном свете | В любое время |
Где протекает | Клетки, содержащие хлорофилл | Все живые клетки |
Кислород | Выделение | Поглощение |
СО2 | Поглощение | Выделение |
Органические вещества | Синтез, частичное расщепление | Только расщепление |
Энергия | Поглощается | Выделяется |
Вот чем отличается от дыхания фотосинтез. Таблица наглядно показывает основные их различия.
Некоторые «парадоксы»
Большая часть дальнейших реакций протекает тут же, в строме хлоропласта. Дальнейший путь синтезированных веществ различен. Так, простые сахара сразу выходят за пределы органоида, накапливаясь в других частях клетки в виде полисахаров, прежде всего — крахмала. В хлоропластах происходит как отложение жиров, так и предварительное накопление их предшественников, которые затем выводятся в другие области клетки.
Следует четко понимать, что все реакции синтеза требуют колоссального количества энергии. Единственным ее источником является все тот же фотосинтез. Это процесс, который зачастую требует столько энергии, что ее приходится получать, разрушая вещества, образованные в результате предыдущего синтеза! Таким образом, большая часть энергии, которая получается в его ходе, затрачивается на проведение множества химических реакций внутри самой растительной клетки.
 Лишь некоторая ее доля используется для непосредственного получения тех органических веществ, которые растение берет для собственного роста и развития либо откладывает в форме жиров или углеводов.
Лишь некоторая ее доля используется для непосредственного получения тех органических веществ, которые растение берет для собственного роста и развития либо откладывает в форме жиров или углеводов.Статичны ли хлоропласты?
Принято считать, что клеточные органоиды, в том числе и хлоропласты (строение и функции которых нами подробно расписаны), находятся строго в одном месте. Это не так. Хлоропласты могут перемещаться по клетке. Так, на слабом свету они стремятся занять положение близ наиболее освещенной стороны клетки, в условиях средней и слабой освещенности могут выбирать некие промежуточные положения, при которых удается «поймать» больше всего солнечного света. Это явление получило название «фототаксис».
Как и митохондрии, хлоропласты являются довольно-таки автономными органоидами. У них имеются собственные рибосомы, они синтезируют ряд высокоспецифичных белков, которые используются только ими. Есть даже специфичные ферментные комплексы, при работе которых вырабатываются особые липиды, требуемые для построения оболочек ламелл. Мы уже говорили о прокариотическом происхождении этих органоидов, но следует добавить, что некоторые ученые считают хлоропласты давними потомками каких-то паразитических организмов, которые сперва стали симбионтами, а затем и вовсе превратились в неотъемлемую часть клетки.
Значение хлоропластов
Для растений оно очевидно – это синтез энергии и веществ, которые используются растительными клетками. Но фотосинтез — это процесс, который обеспечивает постоянное накопление органического вещества в масштабах всей планеты. Из углекислого газа, воды и солнечного света хлоропласты могут синтезировать огромное количество сложнейших высокомолекулярных соединений. Эта способность характерна только для них, и человек пока далек от повторения этого процесса в искусственных условиях.
 Вся биомасса на поверхности нашей планеты обязана своим существованием этим мельчайшим органоидам, которые находятся в глубинах растительных клеток. Без них, без проводимого ими процесса фотосинтеза на Земле не было бы жизни в ее современных проявлениях.
Вся биомасса на поверхности нашей планеты обязана своим существованием этим мельчайшим органоидам, которые находятся в глубинах растительных клеток. Без них, без проводимого ими процесса фотосинтеза на Земле не было бы жизни в ее современных проявлениях.Надеемся, вы узнали из этой статьи о том, что такое хлоропласт и какова его роль в растительном организме.
fb.ru